This is a protocol for the determination of oxygen consumption rate (OCR) in living C. elegans using the Seahorse apparatus with 24-well plates. OCR provides a measurement of respiration, which can be altered by genetic or pharmacologic manipulation of the mitochondrial respiratory chain. Although the Seahorse was originally designed for use with ex vivo cells, it can readily be used in vivo with living C. elegans by following our simple procedure,2 described here for the analysis of four samples side-by-side.
Synchronize each of the four samples of worms. We allow 200 adults to lay eggs for 4 h on a 10 cm plate by picking 200 adults to a 10 cm plate, kill the adults, then study the F1s as day 1 adults. Program the Seahorse Bioscience XF24 Extracellular Flux Analyzer with the following protocol: 1- calibrate probes; 2- loop 10 times; 3- mix 2 min; 4- time delay 2 min; 5- measure 2 min; 6- loop end. Furthermore, the internal heater should be switched off to allow the instrument to be close to room temperature. Hydrate the probes in the extracellular flux assay kit by adding 1 ml of Seahorse calibrant solution to each well and incubating overnight at 37 oC . Load this into the Seahorse on the day of the experiment for calibration.
Rinse the worms off the plate and wash three times using gravity separation in M9 and resuspend each washed sample in 2 ml. Get a rough estimate of the concentration of worms by pipetting 5 drops of 20 µl each per sample onto an empty dish and counting under the dissecting scope. There should be about 2 worms per drop. In the Seahorse assay plate, place 500 µl M9 into four blank wells. At least 30 min after the worms were first rinsed, pipet the four worm samples into a total of 500 ul M9 in each of the other wells with between 25 and 150 worms per well and five replicates of each sample. We use a low retention tip cut to create a wider opening, and found that picking worms into the wells with a platinum wire leads to unstable results.
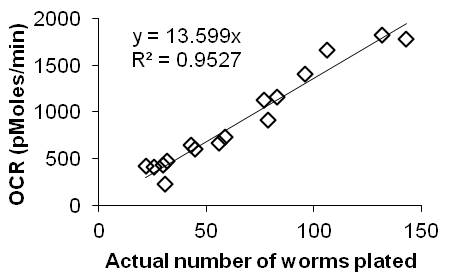
When prompted by the instrument, replace the calibration plate with the worm plate. At the end of the run, count the actual number of worms per well; for this purpose, we capture images on a camera-fitted dissecting scope. Average the background-corrected OCR (pMol O2/min) across the 10 time-points for each well, then normalize the OCR to the actual number of worms in each well. Calculate the average and standard deviation of OCR per worm across the five replicates for each sample. A moderate amount of variation is expected between time-points, wells, plates, and days: we therefore use a minimum of 10 time points of 5 wells, and on at least 2 plates and/or days, to get a meaningful result.
1 For a more detailed protocol with additional tips, please contact Beverley at Beverley.M.Dancy@gmail.com.
2 We acknowledge other labs using Seahorse for C. elegans who provided some tips when we began this work, including Laurent Mouchiroud in Johan Auwerx’s lab at Ecole Polytechnique Fédérale de Lausanne, David Raizen at University of Pennsylvania, and Paul Brookes at University of Rochester Medical Center.
3 We thank José Antonio Mari (ja_mari@yahoo.com) for putting together the graphics in Figure 1.
Figures

A modified leaving assay to study pathogen avoidance
C. elegans constantly encounter pathogenic threats and the ability to recognize and avoid these threats is crucial to survival. C. elegans have evolved a variety of immediate and learned behavioral responses to minimize pathogen exposure (Yook and Hodgkin, 2007; McMullan et al., 2012; and others). Behavioral responses to pathogens are commonly investigated using a food choice assay where animals are given a choice between their standard laboratory food, OP50, and the pathogen of interest (Yook and Hodgkin, 2007; McMullan et al. 2012). The results of this type of assay integrate both the known attractive effect of OP50 and the attractive/repellent effect of the pathogen under investigation. In order to examine only potential repellent cues, we have adapted the male leaving assay (Lipton et al., 2004) to analyze the response of naïve animals upon exposure to Microbacterium nematophilum, a gram positive bacteria which binds to and colonizes the rectal opening of C. elegans.
Method: 90 mm NGM assay plates were prepared using 13 ml NGM in order to make animals tracks easier to observe. Bacterial cultures were grown to OD600 = 1.0 before plating 20 µl in the center of the assay plate.
Single synchronized young adult hermaphrodites were transferred onto individual plates. 10 plates were prepared for each bacterial lawn to be tested.
Plates were kept at 20 oC and examined at 1 hr intervals to determine if the animal was a leaver, meaning that the animal or its tracks can been seen 1 cm or less from the edge of the plate. Animals determined to have left were not scored again at later time points
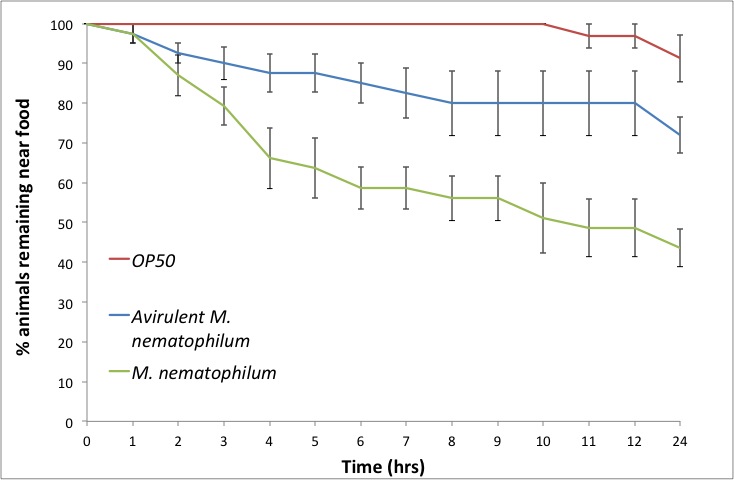
Results: Animals placed on plates seeded with OP50 show little leaving behaviour over 24 hrs. An avirulent M. nematophilum strain (Akimkina et al., 2006) induces a limited leaving response, whilst virulent M. nematophilum induces a rapid response over 24 hrs (Figure 1).
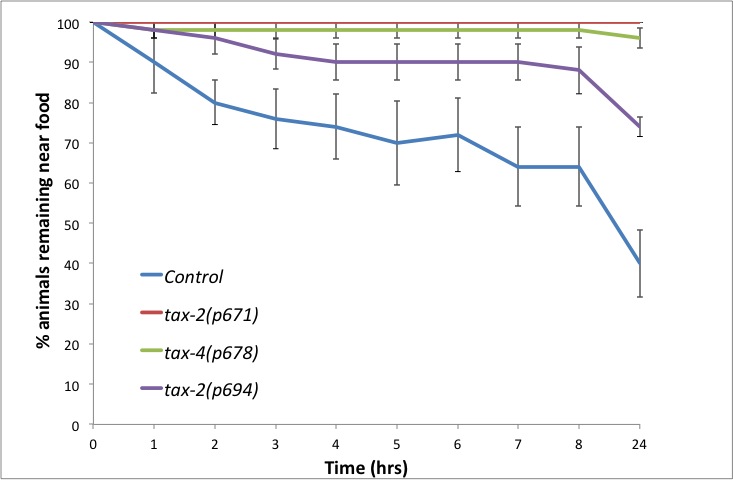
Using this assay we tested the role of the cyclic nucleotide gated ion channel TAX-2/TAX-4 in this behaviour. This pathway has been shown to be required for wild type responses to M. nematophilum using a food choice assay (Yook and Hodgkin 2007). Avoidance of M. nematophilum depends on TAX-2/TAX-4, since mutations in either TAX-2 or TAX-4 abrogate the leaving response (Figure 2). In wild-type animals TAX-2/TAX-4 is expressed in AWC, AFD, ASE, ASG, ASJ, AQR, BAG, ASK, ASI, AWB, and PQR (Coburn and Bargmann, 1996). The allele tax-2(p694) has a deletion in the promoter region and first exon of tax-2 abolishing its expression in ASE, AQR, AFD, and BAG (Coburn and Bargmann, 1996). In contrast to the response to M. nematophilum in a food choice assay (Yook and Hodgkin, 2007), tax-2(p694) animals show a defective leaving response particularly at longer time points (Figure 2), suggesting these four cells are important for the normal leaving response to this pathogen.
This simple behavioural assay could be adapted to investigate the response to numerous pathogens and removes the influence of the attractive cue from OP50 in food choice assays.
Acknowledgements: We thank Arantza Barrios (University College London) and Richard Poole (University College London) for suggestions and assistance in setting up this assay. We thank Jonathan Hodgkin for M. nematophilum strains.
Figures
References
Akimkina T, Yook K, Curnock S, and Hodgkin J. (2006). Genome characterization, analysis of virulence and transformation of Microbacterium nematophilum, a coryneform pathogen of the nematode Caenorhabditis elegans. FEMS Microbiol. Lett. 264, 145–151. 
Coburn CM, and Bargmann CI. (1996). A putative cyclic nucleotide-gated channel is required for sensory development and function in C. elegans. Neuron 17, 695–706.
Lipton J, Kleemann G, Ghosh R, Lints R, and Emmons SW. (2004). Mate searching in Caenorhabditis elegans: a genetic model for sex drive in a simple invertebrate. J. Neurosci. 24, 7427-7434.
McMullan R, Anderson A, and Nurrish S. (2012). Behavioral and immune responses to infection require Gαq-RhoA signaling in C. elegans. PLoS Pathog 8, e1002530.
Yook, K. and Hodgkin J. (2007). Mos1 mutagenesis reveals a diversity of mechanisms affecting response of Caenorhabditis elegans to the bacterial pathogen Microbacterium nematophilum. Genetics 175, 681-697.
A simple method for quantifying M. nematophilum clearance from the rectal opening of C. elegans
In its natural environment C. elegans encounters many bacterial pathogens and is capable of mounting a conserved immune response to protect itself from these infections. Microbacterium nematophilum (M. nematophilum) is a coryneform gram positive bacteria which binds to the post anal cuticle and colonises the rectal opening of C. elegans. This leads to a chronic, non-lethal infection characterised by gut distension, constipation and an innate immune response that includes swelling of the post-anal region known as the deformed anal region (DAR) phenotype (Hodgkin et al., 2000). M. nematophilum binding to the rectal opening can be visualised using the nucleic acid stain SYTO13 (Invitrogen), which shows green fluorescence when excited at 488-491 nm (Figure 1). Several groups including our own have used this nucleic acid stain to detect the presence of M. nematophilum colonisation in the rectal opening of animals that display the bacterially unswollen (BUS) phenotype following infection (Figure 1) (Nicholas and Hodgkin, 2004; McMullan et al., 2012).
An important mechanism of the host defence system to combat and resolve infections is the ability to clear pathogens. We have utilised the easy detection of SYTO13 to follow the ability of C. elegans to clear M. nematophilum from its rectal opening. By scoring the number of animals in a population that show green fluorescence in the rectal opening at 30 minute time points we can see a decrease in M. nematophilum adherence over time (Figure 2). This reflects loss of M. nematophilum attachment rather than fluorescent decay or loss only of the SYTO13 stain, as re-incubation of the animals with fresh SYTO stain at a 90 minute time point results in the same number of animals with green fluorescence.
Bus mutants which still show pathogen binding, such as mutants in the EGL-30–UNC-73–RHO-1 pathway (McMullan et al., 2012) show a corresponding decrease in their ability to clear the SYTO13 positive M. nematophilum from the rectal opening (Figure 2).
This is the first demonstration of the positive role of the DAR phenotype in aiding pathogen clearance during infection and confirms that the DAR phenotype acts as part of the protective response. This method of quantifying M. nematophilum clearance could be applied to other pathogens where GFP labelled variants cannot be generated making it a simple method to assay the effect of mutations on an animal’s ability to clear infection.
Method: Adult animals were transferred from OP50 plates to infection plates seeded with 10% M. nematophilum and the progeny grown to L4-adult stage.
SYTO13 labeling was performed as described previously (Nicholas and Hodgkin, 2004) except that after a 60 minute incubation with SYTO13 10-20 µl of settled, stained worms were transferred to NGM plates lacking food. A ring of 150 mM Copper Sulphate, 2% SDS was used to prevent animals escaping from the plates.
The number of animals colonized by SYTO13 positive M. nematophilum was scored using a Nikon SMZ1500 microscope with GFP filter.
Acknowledgements:We thank Stephen Nurrish (University College London) for suggestions in setting up this assay. We thank Jonathan Hodgkin for M. nematophilum strains.
Figures
References
Hodgkin J, Kuwabara P, and Corneliussen B. (2000). A novel bacterial pathogen, Microbacterium nematophilum, induces morphological change in the nematode C. elegans. Curr. Biol. 10, 1615–1618.
McMullan R, Anderson A, and Nurrish S. (2012). Behavioral and immune responses to infection require Gαq-RhoA signaling in C. elegans. PLoS Pathog 8, e1002530.
Nicholas H, and Hodgkin J. (2004). The ERK MAP kinase cascade mediates tail swelling and a protective response to rectal infection in C. elegans. Curr. Biol. 14, 1256–1317.
Alzheimer’s disease and gender differences in C. elegans
The prevalence and severity of Alzheimer’s disease (AD) are influenced by gender such that two thirds of AD patients are women. However, the biological mechanisms underlying these sex differences are not fully understood. AD begins with slight memory loss and confusion, and eventually leads to severe mental impairment (Mc Khann et al., 1984). It has been previously shown by Guillozet et al. that the formation of amyloid plaques and neurofibrillary tangles in the brain are responsible for memory and other cognitive functions (Guillozet et al., 2003). A recent human trial of AD treatments has proven to be largely unsuccessful. A probable reason is that multiple factors such as food, social relationships, physical activity, gender, climate and stress all play a role in its pathology. One of the most important factors is gender (Viña and Lloret, 2010). Lack of biological gender model systems for AD limits our understanding of the role that gender play in AD pathogenesis. Elucidating the role of gender in an AD model is crucial to prevention, diagnosis, and treatment of AD in different sexes.
In order to examine the role of gender on the etiology of AD, a C. elegans model was used to emulate the pathological processes of AD. The worm apl-1 gene encodes two identical isoforms, orthologous to the human amyloid precursor protein (APP) involved in AD (Wentzell and Kretzschmar, 2010). Recently, a transgenic animal (CL4176) expressing Aβ42 in muscle tissues (Drake et al., 2003) has been established as a model. Interestingly, the Aβ protein expressed in C. elegans self-aggregates like Aβ1–42, and forms fibrillar structures (Link, 1995).
We used male and hermaphrodite CL4176 transgenic animals and performed a paralysis assay (Drake et al., 2003). Synchronized eggs were maintained at 16 ºC and males and hermaphrodites were sorted at late L4 stage. Sorted worms were subjected to 25 ºC to induce the transgene expression and the paralysis was scored successively at 2 hour intervals. The hermaphrodite animals paralyzed earlier in comparison to the males. The mean time duration at which 50% worms were paralyzed was 34.6 hours and 27.3 hours, for males and hermaphrodites, respectively (p< 0.001) (Figure 1A, 1B). The data shows that the male CL4176 takes significantly more time to undergo paralysis compared to the hermaphrodites.
In this paper, we provide a way to study the mechanism and treatment strategy of AD disease based on gender. The observed phenotype is consistent with human beings (Viña and Lloret, 2010). Further studies will allow us to investigate the molecular pathways involved in AD in relation to sex and significantly impact our understanding to identify novel therapeutics in the future.
Figures

References
Drake J, Link CD, and Butterfield DA. (2003). Oxidative stress precedes fibrillar deposition of Alzheimer’s disease amyloid beta-peptide (1-42) in a transgenic Caenorhabditis elegans model. Neurobiol. Aging 24, 415-420.
Guillozet AL, Weintraub S, Mash DC, and Mesulam MM. (2003). Neurofibrillary tangles, amyloid, and memory in aging and mild cognitive impairment. Arch. Neurol. 60, 729-736.
Link, CD. (1995). Expression of human beta-amyloid peptide in transgenic Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A. 92, 9368–9372.
McKhann G, Drachman D, Folstein M, Katzman R, Price D, and Stadlan EM. (1984). Clinical diagnosis of Alzheimer’s disease: report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease, Neurology 34, 939-944.
Viña J, and Lloret A. (2010). Why women have more Alzheimer’s disease than men: gender and mitochondrial toxicity of amyloid-beta peptide. J. Alzheimers Dis. S527- 533.
Wentzell J, and Kretzschmar D. (2010). Alzheimer’s disease and tauopathy studies in flies and worms. Neurobiol. Dis. 40, 21–28.
L1 arrest affects dauer decision in C. elegans
During larval development C. elegans has two distinct time points where it can arrest development until environmental conditions become more favorable. The first one is L1 arrest, which occurs when worms hatch in the absence of food. The second decision point is later, around L1-L2 molt, and if conditions are poor at this time (little food, crowding, and high temperature) worms gain the potential to become dauers. L1 arrest and dauer formation are not mutually exclusive and, in principle, a worm can go through both of them sequentially. However, it is not known whether L1 arrest and dauer formation are coupled, i.e., if experiencing L1 arrest affects a worm’s decision to become a dauer. A naïve prediction would be that worms that experienced L1 starvation should be more susceptible to become dauers since they were already exposed to a harsh environment.
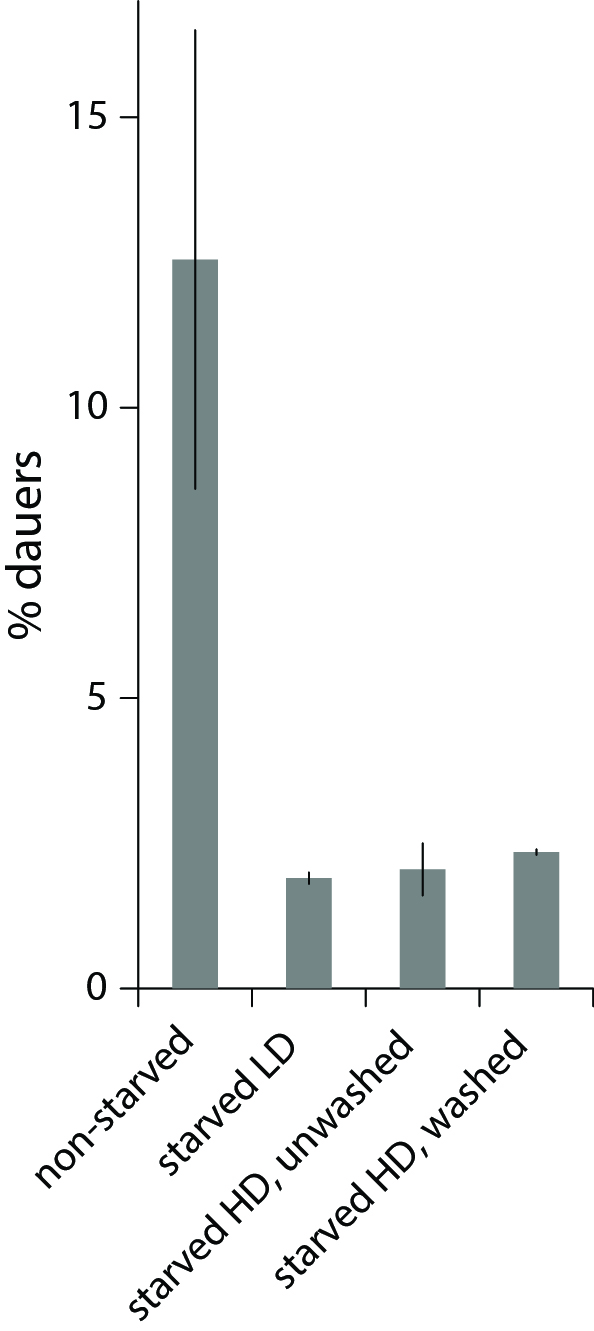
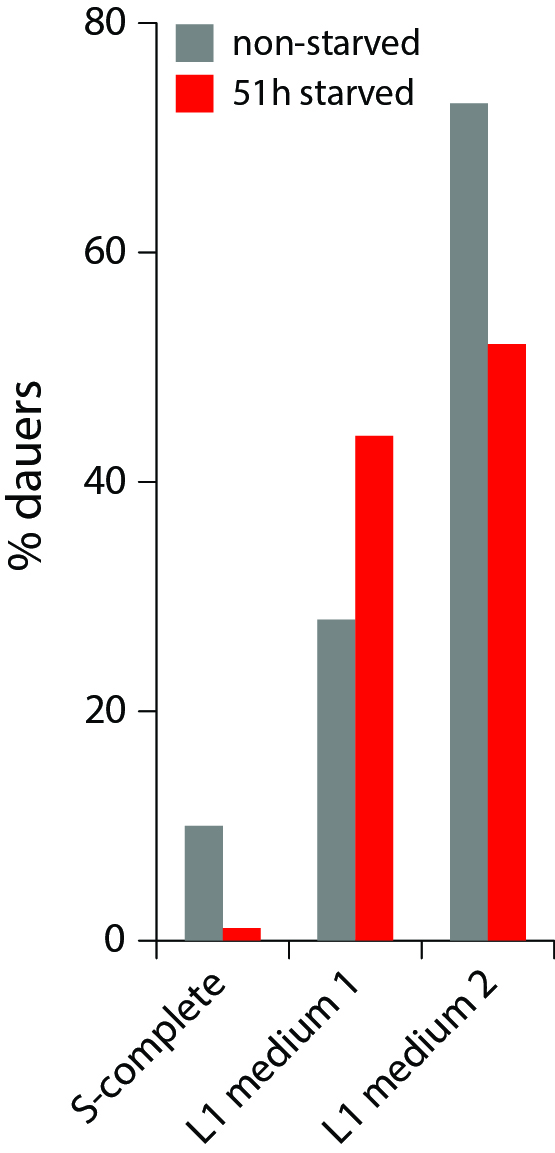
We studied dauer formation of N2 worms in liquid and found that in reality worms that experienced L1 arrest and starvation were less likely to become dauers. In a typical experiment, eggs obtained from an egg prep were incubated at 20 0C in S-complete with a small amount of bacterial food (HB101) and dauer formation was assessed 6 days later based on survival in 1% SDS (Pungaliya et al., 2009). When we let eggs hatch without food and added it 1-4 days later, keeping other conditions the same, significantly fewer worms became dauers (Figure 1). This was not due to slower development after starvation and incomplete dauer formation. We also found that this effect reflected an internal state of the worm rather than exposure to L1 exometabolome during L1 arrest since worms starved as L1s at low and high density later formed dauers with the same probability (Figure 1, during the dauer formation worms were at the same low density of 1 worm/µl to minimize the effect of released chemicals, see below).
Starved L1 worms release numerous metabolites, including several ascarosides, and we anticipated that they might promote dauer formation. Indeed, when S-complete was substituted with conditioned medium from high-density L1 larvae starved for 24 h, more worms became dauers (Fig. 2), consistent with the known dauer-inducing effect of crowding mediated by ascarosides (Ludewig and Schroeder, 2013). However, this result apparently contradicts the starvation effect described above. On one hand, when a worm itself experienced L1 arrest but received no signal from other worms (low density condition), it was less likely to become a dauer and preferred to try to grow to adult. On the other hand, a worm that had not experienced starvation itself but was exposed to chemical signals released by other starved L1 worms, was more likely to become a dauer. The next obvious experiment is to combine these two opposite effects and see which one is stronger. The result of this test (Fig. 2) is that the external starvation signal (mediated by L1 conditioned medium) overrides the internal one (L1 arrest). When making a decision whether to become a dauer, apparently C. elegans worms trust the communal voice more than their own.
While the dauer-inducing effect of L1-conditioned medium is intuitively easy to accept, the negative effect of L1 starvation on subsequent dauer formation is less clear. A recently published model of contrast effects (McNamara et al., 2013) may provide a theoretical framework for understanding this phenomenon. But to keep us from being too comfortable with our simplistic views nature presented us with another puzzle: L1 arrest does not negatively affect dauer formation in C. briggsae.
Figures
References
Ludewig AH and Schroeder FC. (2013). Ascaroside signaling in C. elegans (January 18, 2013), WormBook, ed. The C. elegans Research Community.
McNamara JM, Fawcett TW, and Houston AI. (2013). An adaptive response to uncertainty generates positive and negative contrast effects. Science 340, 1084-1086.
Pungaliya C, Srinivasan J, Fox BW, Malik RU, Ludewig AH, Sternberg PW, and Schroeder FC. (2009). A shortcut to identifying small molecule signals that regulate behavior and development in Caenorhabditis elegans. Proc. Natl Acad. Sci. U. S. A. 106, 7708-7713.