Correspondence to: Greg Nelson (gnelson@dominion.llumc.edu)

View the full size version.
View the full size version.
To perform the procedure, nematodes cleansed of bacteria are picked and then released into a PEG/glycerol droplet (6.5 μl) on a glass slide. A cover slip is then gently placed on the droplet containing the nematode. Due to the small volume of PEG/glycerol droplet, nematodes need to be transferred to PEG/glycerol droplet, and rapidly overlaid with a cover slip before mounting medium (PEG/glycerol) dries. This quick step instantly immobilizes the nematode and leaves them well suited for microcopy, including acquiring images for extended periods of time. We find the method is particularly useful for quick screening of GFP expressing lines, which frequently requires analysis of many nematodes, and use of high-resolution, compound microscopy for detecting low levels of GFP expression (such as due to certain weak promoters).
We have also experimented with other mixtures of PEG and glycerol. For instance, a mixture of 30% PEG 8000 in 25% glycerol also renders the nematodes amenable for microcopy, however increasing PEG concentrations above 30% renders nematodes unstable and unsuitable for microscopy. Larval nematodes appear to be slightly more sensitive to the procedure. We noticed slight cuticle distortion in younger nematodes (L1-L3) when PEG/Glycerol (20:20) solution was used for microscopy. A further dilution of PEG might be required to overcome the distortion and 1 μl of levamisole (20mM stock) can be used to insure that the nematodes are motionless.

Developmentally arrested dauer stages in nematodes for laboratory assays have so far been obtained either by simply starving culture plates or by liquid culture in large Erlenmeyer flasks. Harvesting dauers from starved agar plates is straightforward but the yield is usually very low. Liquid cultures on the other hand produce high yields, but their maintenance is labor-intensive. We have combined the simple setup of starved plates with the increased yield of liquid cultures in the “wet plate protocol”:
It is important to maintain a layer of liquid on the plates at all times. If the worms are able to crawl instead of swim, the plates are already too dry. Just like liquid cultures in an Erlenmeyer flask, liquid plates may become contaminated. It is therefore important to prepare the plates to be as sterile as possible (i.e. under a flowhood). If problems persist, the culture may either be started with freshly bleached eggs instead of a mixed culture. Additionally, antibiotics and fungicides may be added to the worm/OP50 mixture.
For large-scale experiments that require huge numbers of dauers (e.g. microarrays), the use of liquid cultures is preferable. For smaller experiments however, the wet plate protocol has proven to be simpler, quicker and less prone to contamination in many assays in our lab.
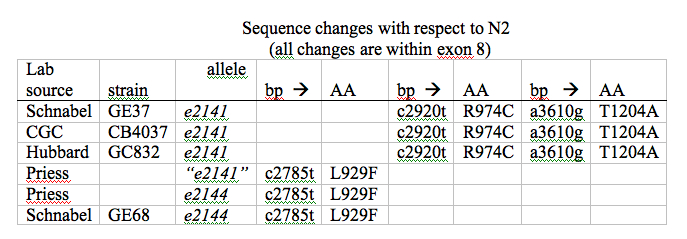
We urge the community to sequence their “e2141“-bearing lab strains prior to future publication or dissemination. We note that in the prior literature, strains designated as “e2141” may have been carrying the e2144 (c2785t) mutation. In light of our sequencing results, the literature indicates that the e2144 (c2785t mutation variously attributed to “e2141” or “e2144“) results in a much stronger embryonic phenotype than e2141 (c2920t a3610g mutations). See, for example, Mello et al., 1994, and Hutter and Schnabel, 1994. The e2141 allele is in the CGC collection as strain CB4037, and the e2144 allele is now deposited at the CGC as strain GE68.
Hutter H and Schnabel R. (1994). glp-1 and inductions establishing embryonic axes in C. elegans. Development 120, 2051-64. 
Kodoyianni V, Maine EM, and Kimble J. (1992). Molecular basis of loss-of-function mutations in the glp-1 gene of Caenorhabditis elegans. Mol. Biol. Cell 3, 1199-213.
Mello CC, Draper BW, and Priess JR. (1994). The maternal genes apx-1 and glp-1 and establishment of dorsal-ventral polarity in the early C. elegans embryo. Cell 77, 95-106.
{kind=link}