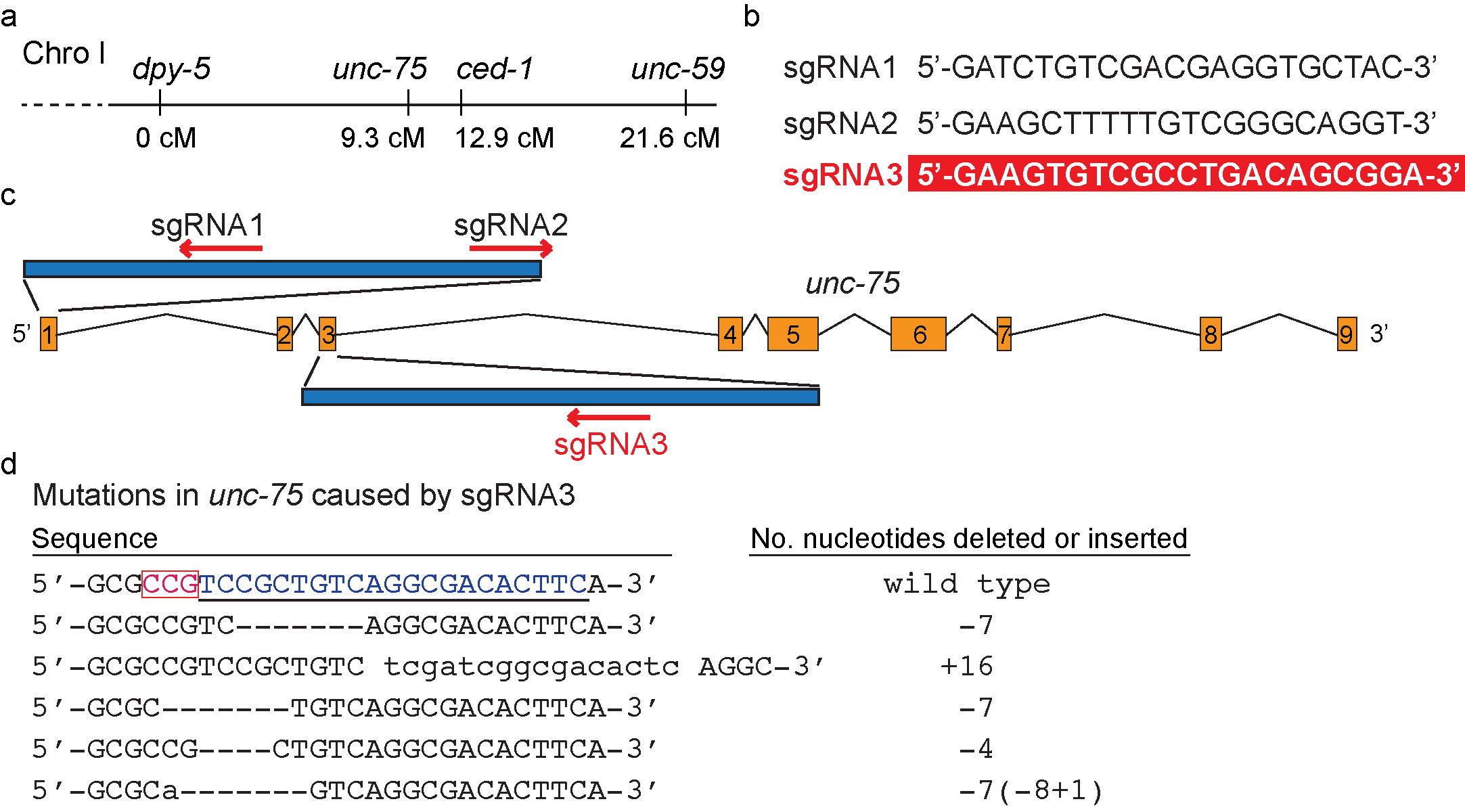
Genetic analysis of mutations that result in phenotypes that are relatively hard to score (weak or partially penetrant growth defects, phenotypes that are only visible under the Nomarski DIC microscope, synthetic phenotypes, etc.) heavily relies on the usage of closely linked, easy-to-score mutations. The traditional method of generating marked mutant alleles is through homologous recombination. Loss-of-function mutations of ced-1, which encodes a C. elegans phagocytic receptor essential for the removal of apoptotic cells (Zhou et al., 2001), result in a cell-corpse persistent phenotype only detectable under the Nomarski DIC microscope. Loss-of-function mutations of unc-75 (Figure 1a) (Loria et al., 2003) cause an Uncoordinated (Unc) phenotype easily detectable under the dissection microscope and are commonly used as linked markers for ced-1. We found that CRISPR/Cas9-mediated knocking out of unc-75 in a ced-1 mutant background is both labor- and time-saving comparing to the traditional method. Here we report our method, result, and the sequence of a small guide RNA (sgRNA) that effectively targets unc-75.We designed three sgRNAs (Figure 1b) predicted to target different regions of unc-75 genomic DNA (Figure 1c) with the help of the online resource (http://crispr.mit.edu/). To express these sgRNA in C. elegans, we cloned each one into plasmid pMB70 (PU6 sgRNA) (Waaijers S et al., 2013). We coinjected all three sgRNA expression constructs (50 ng/µl) together with the Cas9-expression construct pDD162 (50 ng/µl) (Dickinson et al., 2013) and Prab-3 mRFP1 (10 ng/µl) (Mangahas et al., 2008), a transgenic marker, into a ced-1 mutant strain. We found 10 Unc progeny among F1 progeny of 15 injected P0s. Among the progeny of these Unc F1s, we identified three homozygous Unc strains. They did not carry the mRFP1 marker. Sequencing results revealed mutations (Figure 1d) solely in exon 3, within the region targeted by sgRNA3, and no mutations in the regions targeted by sgRNA1 and sgRNA2. Using the same strategy, we isolated additional unc-75 mutant alleles in two other ced-1 mutant backgrounds, in efficiency similar to the above. These alleles also carry mutations solely in the region targeted by sgRNA3 (Figure 1d). In addition, including only sgRNA3 but no sgRNA1 or 2 in the injection mix also resulted in the occurrence, at the similar frequency, of Unc mutations in the F1 generation. The unc-75 mutations we identified include deletions and insertions of up to 16 nucleotides (Figure 1d). Our results thus indicate that sgRNA3 is effective in targeting unc-75 for gene editing, whereas sgRNA1 or sgRNA2 are not.
The frequent occurrence of the Unc-75 mutant worms in the F1 generation indicates that the efficiency of CRISPR/Cas9-mediated gene-editing is high enough to create unc-75 mutations in both homologous alleles. Isolation of Unc mutants in the F1 generation significantly reduces the number of worms to screen through and the time it takes to isolate homozygous mutants. Our observation thus suggests that the CRISPR/Cas9-mediated production of easy-to-score marker mutations is an effective and rapid strategy to generate linked markers that label hard-to-score mutations. We suggest that building a collection of sgRNAs proven successful in targeting commonly used marker genes will benefit the C. elegans community.
Figures
References
Dickinson DJ, Ward JD, Reiner DJ, Goldstein B. (2013). Engineering the Caenorhabditis elegans genome using Cas9-triggered homologous recombination. Nat. Meth. 10, 1028-1034.

Loria PM, Duke A, Rand JB, Hobert O. (2003). Two neuronal, nuclear-localized RNA binding proteins involved in synaptic transmission. Curr. Biol. 13, 1317-1323.
Mangahas PM, Yu X, Miller KG, Zhou Z. (2008). The small GTPase Rab2 functions in the removal of apoptotic cells in Caenorhabditis elegans. J. Cell Biol. 180, 357-373.
Waaijers S, Portegijs V, Kerver J, Lemmens BB, Tijsterman M, et al. (2013). CRISPR/Cas9-targeted mutagenesis in Caenorhabditis elegans. Genetics 195, 1187-1191.
Zhou Z, Hartwieg E, Horvitz HR. (2001). CED-1 is a transmembrane receptor that mediates cell corpse engulfment in C. elegans. Cell 104, 43-56.