To ensure that most, if not all, of the E. coli were removed from worms they were each placed in a drop of M9 buffer for 5 minutes on a blank NGM plate then placed out of the drop on the plate for an additional 5 minutes prior to the trial. Following washing, single worms were placed on a fresh NGM agar plate for an initial 30 minute observation period and then either lifted with a platinum pick and put back onto their original plate (sham switch), lifted and moved to an unexplored NGM plate, or lifted and moved to an NGM plate previously explored by a single conspecific for 30 min. The time the worms were lifted off the plates was held constant between the groups.
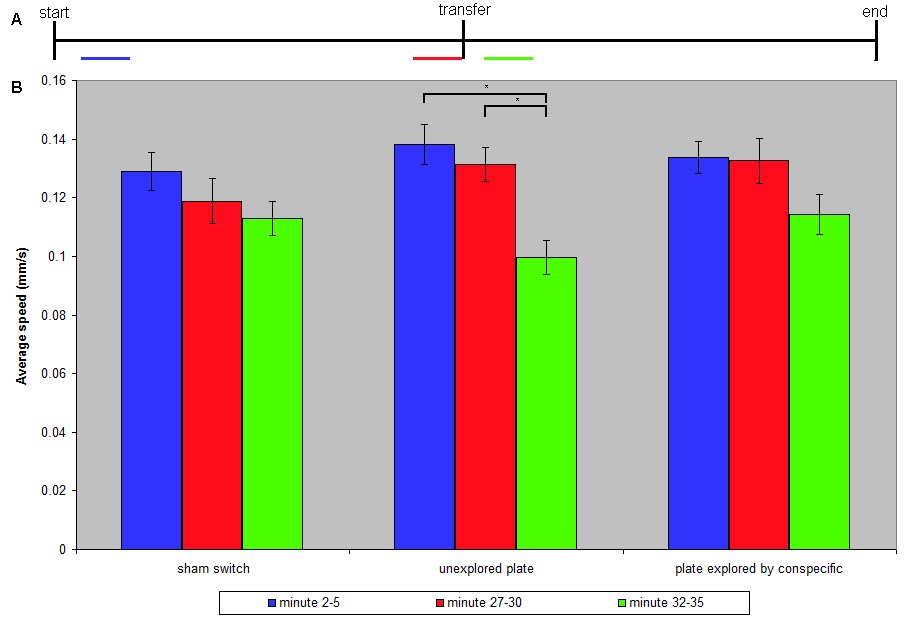
We measured and compared the speed of the worms at the beginning (min 2-5) and end (min 27-30) of the 30 min exploration of the first plate and at the beginning of the time on the subsequent plate (min 32-35). Speed was measured as a proxy for foraging behavior (Bargmann, 2006). Rate of spontaneous reversals and foraging range were found to be insufficiently robust to be suitable measures. For statistical analyses ANOVAs were used to compare the speeds of locomotion between the beginning of the first 30 min (blue bars) to the average speed at end of the first 30 min (red bars), and the average speed at min 32-35 (min 2-5 of the second 30 min period; green bars) for each of the three groups. A Bonferroni correction for multiple tests set significance at p< 0.016.
The first minute after each transfer (to the initial plate and to the new plate after 30 min) all worms moved rapidly (data not shown), therefore the first two minutes of each 30 min period were not included in our analysis. There was no change in the rate of speed from min 2-5 to min 27-30 of the first 30 min observation period for any of the three conditions (Fig 1). ANOVAs for the sham switch group and for the group switched to a plate previously explored by another worm showed no significant differences in speed between the three timepoints tested. In contrast, an ANOVA for the group switched to a blank unexplored plate was significant (p<0.000044): post-hoc tests indicated that the their speed in min 32-35 (min 2-5 of the second observation period) was significantly slower than their speed at either min 2-5 or min 27-30 of the first observation period. Thus C. elegans moved significantly more slowly on a plate that had not been explored by a worm (either themselves or a conspecific) than on a previously explored plate.
Although it is possible that the explored plates might have had trace amounts of E. coli it is less likely than the hypothesis that some worm pheromone allowed worms to distinguish plates that had contained worms from plates that had not. Worm tracks from each of the three groups were very similar and did not indicate that worms on a blank plate explored any differently that worms in the two “previously explored plate” conditions. Unexplored plates elicited slower movement than plates containing worm odor. Taken together the data suggest that C. elegans do not have the ability to distinguish self from conspecifics (and pilot data suggests self from C. briggsae), but do distinguish between environments where worms have been and environments where there have been no worms.
Figures
References
Bargmann, C. I. (2006). Chemosensation in C. elegans, Wormbook, ed. The C. elegans Research Community, Wormbook, http://www.wormbook.org/.