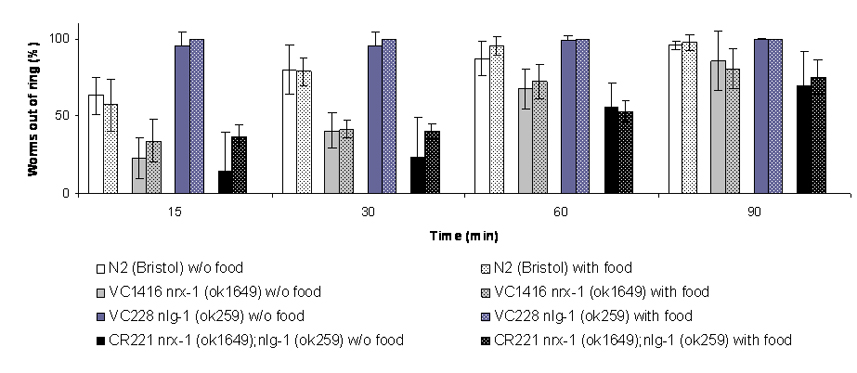
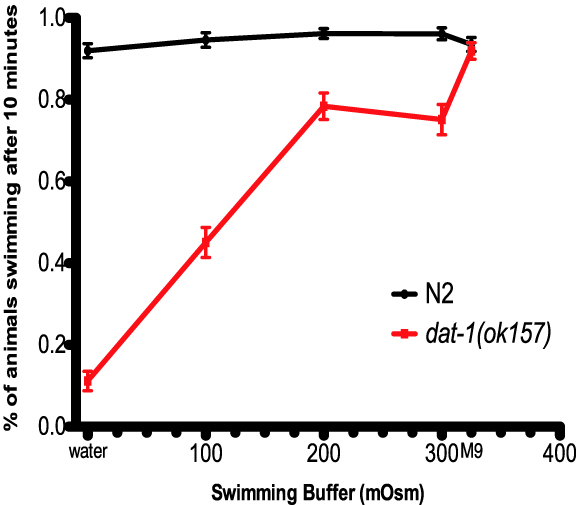
Dopamine (DA) signaling depends upon the coordination of multiple presynaptic processes, including those supporting DA biosynthesis, packaging, release and reuptake. The presynaptic dopamine transporter (DAT-1) is of particular importance in regulating both the exposure of pre- and postsynaptic DA receptors as well as supporting recycling of DA for further rounds of release. Using Caenorhabditis elegans, our lab demonstrated that loss of DAT (dat-1) elicits a DA-dependent, locomotory phenotype termed Swimming Induced Paralysis (SWIP) (Mcdonald et al., 2007). Whereas wild-type (N2) L4 stage worms swim at a continuous rate for ~20 min., dat-1 worms initiate normal swimming, but then paralyze over the next six minutes. SWIP can be rescued by pretreatment with the vesicular DA transporter (CAT-1) inhibitor reserpine, loss of DA biosynthesis (cat-2) or loss of the DA receptor DOP-3 (dop-3). In our initial studies, dat-1 SWIP was thought to be evident in either water or M9 buffer. Recently, however, we tested N2 and dat-1 worms for SWIP in both media, as well as three sucrose-supplemented water solutions. As shown in Fig. 1, as the osmolarity of the aqueous media increases, dat-1 SWIP penetrance decreases significantly. In contrast, swimming of wild-type worms is unaffected by media osmolarity. Although the effects of osmolarity on neural activity in worms have been well documented, here we show that media composition modulates a DA-modulated C. elegans motor behavior by DA and possibly other biogenic amines like serotonin. Candidate, reverse or forward genetic manipulations of SWIP in water vs. M9 may provide an opportunity to further dissect DA signaling in the nematode in vivo.
Figures
References
Mcdonald PW, Hardie SL, Jessen T, Carvelli L, Matthies DS, Blakely RD. (2007). Vigorous motor activity in Caenorhabditis elegans requires efficient clearance of dopamine mediated by synaptic localization of the dopamine transporter DAT-1. J. Neurosci. 27, 14216-14227.