
View the full size version.
View the full size version.
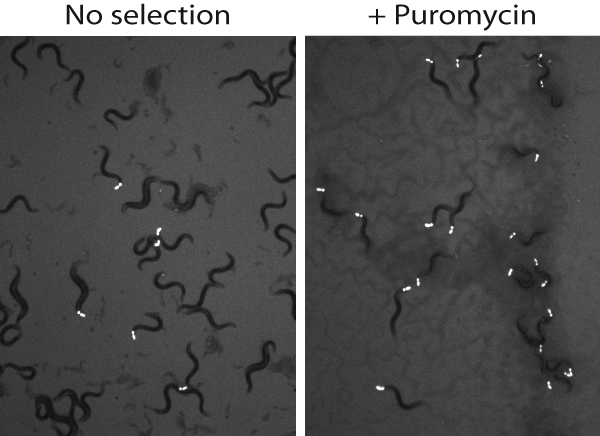
Biolistic transformation is currently a popular method for making trangenic lines in C. elegans. Although this method is reliable, the fraction of successfully transformed animals that is obtained is extremely small, necessitating the use of a co-transformation marker. The most commonly used marker is the unc-119(+) gene which can dominantly rescue the severe Unc and dauer defective phenotypes of unc-119(ed3) mutants. Despite the utility of the unc-119(+) marker, screening for rare transgenic animals amongst the progeny of bombarded animals is time consuming and labor intensive. One approach is to allow the plates containing the bombarded animals to starve over the course of a few weeks. Each plate is then divided into chunks and the chunks moved to fresh plates containing food. These plates can then be screened a day or so later for animals with normal mobility. This approach is still time consuming and has the added drawback of amplifying the number of plates (and therefore the surface area) that has to be scanned. I have devised a simple alternative to this approach in which the worms do most of the work. One to two weeks after bombardment, just after the plates have starved, a fresh plate containing OP50 is cut up into little (1-2 cm2) chunks. Each of these chunks is placed on one of the plates containing the bombarded animals so that the side containing the bacteria is face up (away from the worms). The plates are left upright in the incubator overnight and the next day the bacterial lawn on top of the transferred chuck is checked for rescued worms. Because the animals have to climb up to the top of the chuck to reach the food, the rescued animals have a significant advantage. Using this approach, my group has on multiple occasions, been able to identify a single transformed worm from a 100 mm plate. This approach has the added advantage of being rapid; rather than scouring the entire plate for transgenic worms, only the small area on top of these “pedestals of rescue” has to be searched.
Two of our methods are now seeing publication. We are writing to let people know how to get the relevant strains and constructs. The first method uses the ninth intron of mec-2, whose splicing depends on mec-8, to produce temperature-sensitive expression and temperature-sensitive RNAi (Calixto et al., 2010a). The intron 9 vector TU#821 (Pmec-18 intron-9 mec-2::yfp) is available from Addgene (www.addgene.org/Martin_Chalfie). The TU218 [mec-8(u218ts)] and TU3135 [mec-8(u218); rde-1(ne219); uIs46 (ceh-22::GFP, Prde-1mec-2 intron9::rde-1)] strains are available from the C. elegans Genetic Center (CGC).
The second method uses SID-1 expression in neurons to allow feeding RNAi for neuronally-expressed genes (Calixto et al., 2010b). Expression of neuronal sid-1 in the RNAi sensitized strain lin-15b(n744) greatly enhanced this effect. Furthermore, promoter-driven sid-1(+) can be used with a sid-1 mutant background to restrict RNAi to specific cells. Currently we are using neuronal and touch receptor neuron-specific expression of sid-1 to uncover neuronal and touch neuron-specific defects for genes whose loss has more general effects (e.g. lethality). The following strains have been submitted to the CGC. In addition, a vector allowing pan-neuronal sid-1 expression TU#867 (Punc-119 sid-1) will be available from Addgene.
| Strain name | Genotype |
|---|---|
| TU3311 | uIs60 (Punc-119yfp, Punc-119sid-1) |
| TU3335 | uIs57 (Punc-119yfp, Punc-119sid-1, Pmec-6mec-6); lin-15b(n744) |
| TU3401 | uIs69 [pCFJ90 (Pmyo-2mCherry), Punc-119sid-1]; sid-1(pk3321) |
| TU3595 | uIs72 [pCFJ90 (Pmyo-2mCherry), Punc-119sid-1, Pmec-18mec-18::gfp]; sid-1(pk3321) him-5(e1490); lin-15b(n744) |
| TU3568 | uIs71 [pCFJ90 (Pmyo-2mCherry), Pmec-18sid-1]; sid-1(pk3321) him-5(e1490); lin-15b(n744) |
Calixto A, Ma C, Chalfie M. (2010a). Conditional gene expression and RNAi using MEC-8-dependent splicing in C. elegans. Nat. Methods 7, 407-411. 
Calixto A, Chelur D, Topalidou I, Chen X, Chalfie M. (2010b). Enhanced neuronal RNAi in C. elegansusing SID-1, Nat. Methods, [Epub ahead of print].
The adherence of worms to standard plastic tips makes accurate dispensing difficult. While using glass tips can overcome this problem, their application is limited when sterility and frequent tip changing is needed, for example, when performing liquid culture RNAi screens using 96-well plates (Lehner et al., 2006). RNAi screen protocols often require ~10 L1 worms to be dispensed into each well of a 96 well plate, however using standard P200 tips with worms in M9 alone resulted in an average variance of ± 6 L1 worms per well. We found that worms suspended in M9 with Triton X-100 decreased this adherence, resulting in an average variance of ± 2 L1 worms in each 10µl volume.
Both Triton X-100 and Tween-20 can be used to decrease this adherence to standard P200 tips. Since autoclaving either of these reagents is not recommended, sterilization can be carried out via filtration using a 2µm filter. The lower viscosity of Triton X-100 over Tween-20 greatly aided filtration and sterility tests showed no associated contamination. We found that using concentrations of Triton X in M9 ranging from 0.1% to as low as 0.01% proved to be effective in decreasing worm adherence throughout our assay. No negative issues regarding the growth or progeny of worms were observed when compared to a control plate containing no Triton X. Therefore this method could serve useful when applied to protocols that require consistently small numbers of worms in large repetitions.
Lehner B, Tischler J, Fraser, AG. (2006). RNAi screens in Caenorhabditis elegans in a 96-well liquid format and their application to the systematic identification of genetic interactions. Nat. Protoc. 1, 1617-20.
{kind=link}