
View the full size version.
View the full size version.
Those of us who have worked on C. elegans for many years fondly remember The Worm Breeder’s Gazette (the C. elegans newsletter) and probably still have all the old copies on our shelves. The Gazette was how we let each other know what we were doing, share new techniques, and, more generally, enthuse about this best of all model organisms. On a personal note, the Gazette was where I first announced our results with GFP. For much of its existence virtually everyone contributed to the Gazette. Thus, many of us were sad when the Gazette stopped being published in 2003. I am happy to report that the Gazette did not die; it just took a vacation. Now under the sponsorship of WormBook, the Gazette is back as an online journal that will be published every six months.
The Gazette was started by Bob Edgar at the University of California at Santa Cruz in December, 1975. One of Bob’s goals for the Gazette was that it would unite researchers working on C. elegans. With the Gazette he started the tradition of sharing data that has, for many of us, most characterized our group. As he wrote in his forward to the first issue: “I hope it. . .proves useful to you and stimulates informal interchange of ideas and information.” By 1983, the Gazette moved to the Caenorhabditis Genetics Center at the University of Minnesota, where it was assembled by Bob Herman and Theresa Stiernagle. Later Leon Avery began a web site, so that people could have access to all the Gazette articles online. Another pivotal person in Gazette history is Greg Nelson, who drew many of the covers and (fortunately for us) has agreed to continue providing artwork for the new edition.
The idea to revive the Gazette began at a Janelia Farm meeting on C. elegans neurobiology in March of this year. When asked what would best aid their research, many people said that they wanted a way to keep abreast of new techniques. A rejuvenated Gazette seemed to be a good solution. We announced the return of the Gazette at the Los Angeles Worm Meeting in June, and now have our first issue. Not surprisingly, putting together a newletter, is not an easy job, and if it were not for Jane Mendel, Daniel (Qinghua) Wang, and Todd Harris (the ones who did all the work), the new Gazette would not have been possible.
Because the Gazette is now an online journal, we can do more than was done with the hardcopy version. For example, contributors can include color figures, and links can be provided for additional information in WormBase or other web sites. Readers can now also leave comments. We envision that the Gazette will continue to be a place where C. elegans researchers and researchers studying other nematodes alert their colleague about interesting results, new technique, and observations that can make their research better. It is a pleasure to say that the Gazette is back. Please enjoy it and give us your feedback and suggestions.
A number of gene expression studies done in single cell organisms have shown that variable gene expression due to stochastic molecular processes can lead to phenotypic variability, and because of that genes must be regulated in an appropriate way in each cell. This becomes much more intricate during C. elegans development when multiple cells also have to communicate with one another via various molecular signals susceptible to intrinsic and/or extrinsic fluctuations but still have to divide timely and adopt their proper cell fates.
In the need of quantitative measurement of endogenous gene expression to tackle these problems, our lab has recently developed an in situ hybridization method which allows us to detect individual copies of mRNA molecules in C. elegans embryos and larvae as diffraction-limited fluorescent spots (Raj et al., 2008 ). Using the available nucleic acid sequence of the target mRNA, we designed 48 different singly labeled ~20bp long oligonucleotide probes. Having a large number of probes targeting one transcript ensures the signal-to-noise ratio to be much less sensitive to non-specific binding of the probes and only the target mRNA molecules appear as a bright fluorescent spots in the image.
Our new method has the following advantages: 1) we can measure discrete quantity and location of mRNA molecules in fixed animals with preserved shape, 2) it is quick – custom designed oligonucleotide probes can be commercially synthesized in less than a week, 3) multiple genes can be probed simultaneously – so far we were able to reliably detect up to three different genes by labeling each probe with three different fluorophores in addition to DAPI nuclear staining, and 4) we can measure endogenous mRNA level and avoid artifacts of genetic constructs which often lack unidentified key regulatory regions such as UTRs, intronic sequences, etc.
More details of our in situ hybridization technique including detailed protocols and probe designer algorithm are available online at http://www.singlemoleculefish.com/

Raj A, van den Bogaard P, Rifkin SA, van Oudenaarden A, and Tyagi S. (2008). Imaging individual mRNA molecules using multiple singly labeled probes. Nat. Methods 5, 877-879. 
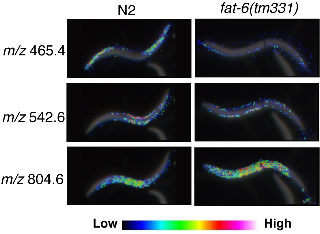
Imaging mass spectrometry (IMS) is a technique based on two-dimensional mass spectrometry to visualize the spatial distribution of biomolecules, which does not need separation, purification or labeling of target molecules (Kimura et al., 2009). Using tissue sections of mammals we, and others, have succeeded to visualize the distribution of peptides, lipids and other small metabolic molecules. Here, we report the first application of IMS to C. elegans. MALDI (matrix-assisted laser desorption ionization) imaging has revealed that numerous biomolecules are localized specifically in the limited regions. The resolutions of the images obtained by IMS are high enough to analyze the biomolecules even in small C. elegans (Fig.).
We compared wild type (N2) and fat-6(tm331) mutants, which lack a Δ9 fatty acyl desaturase, and observed a dramatic difference in expression level and localization of biomolecules (Fig.). We have not studied the molecular identity of these molecules yet, however, MS/MS analysis can give us detailed nature of the molecular structure. We hope that IMS could bring innovation to the C. elegans research community as a new microscopy to observe worms and a convenient tool for mutant screening.
Kimura Y, Tsutsumi K, Sugiura Y and Setou M. (2009). Medical molecular morphology with imaging mass spectrometry. Med. Mol. Morphol. 42, 133-137.
Current methods for in vivo patch-clamp electrophysiology and optical imaging of calcium transients in C. elegans neurons rely on veterinary-grade cyanoacrylate glue for immobilizing animals (Goodman et al., 1998; Lockery and Goodman, 1998; Kerr, et al., 2000). Just about ten years have passed since the first studies measuring cellular responses in individual neurons were published, all of them using Nexaband S/C (WPI, Sarasota, FL) to immobilize worms on thin, 2% agarose pads. Recently and tragically, Abbott discontinued the manufacture of Nexaband S/C. HistoacrylBlue has been used to immobilize animals during recordings from the C. elegans neuromuscular junction (Richmond and Jorgensen, 1999). Unfortunately, the dye added to this adhesive is fluorescent when viewed under a GFP filter set, obscuring GFP-tagged neurons.
Alternative methods for immobilization have emerged from the recent explosion in the design and fabrication of microfluidics devices for worms (Ben-Yakar et al., 2009). These devices have been used for long- and short-term imaging work and appear to be significantly less toxic than even the best cyanoacrylate adhesives. However, the need for physical access to the animals during an electrophysiology experiment precludes the use of existing traps for in vivo recording. Thus, despite the great advances provided by microfluidics devices, there is a continuing need for a minimally toxic, cyanoacrylate adhesive.
We are pleased to report a suitable alternative, custom-formulation from GluStitch Inc. (http://www.glustitch.com, contact: Steven Blacklock, sales@glustich.com): 80% octyl-20% butyl cyanoacrylate (r = 5.75 centiPoise). The viscosity and polymerization rate of this glue is such that it can be applied in a targeted way to one side of a worm without coating the entire animal. With this formulation, it is possible to see the GFP label and also to expose GFP-tagged neurons of interest, the first step needed for electrophysiological recordings. Additionally, worms appear healthy hours after being glued. Other glues and adhesion methods tested were deemed unsuitable. These include: Nexcare (3M), other formulations from GluStitch, Inc. (high viscosity (75 cP) 100% butyl cyanoacrylate, 50/50 octyl/butyl cyanoacrylate, 100% butyl cyanoacrylate (3.7 cP)) and Gluture (Abbott Laboratories). Good luck and good patching.
Ben-Yakar A, Chronis N, and Lu H. (2009). Microfluidics for the analysis of behavior, nerve regeneration and neural cell biology in C. elegans. Curr. Opin. Neurobiol. [Epub ahead of print].
Goodman MB, Hall DH, Avery L, and Lockery SR. (1998). Active currents regulate sensitivity and dynamic range in C. elegans neurons. Neuron 20, 763-772.
Kerr R, Lev-Ram V, Baird G, Vincent P, Tsien RY, and Schafer WR. (2000). Optical imaging of calcium transients in neurons and pharyngeal muscle of C. elegans. Neuron 26: 583-594.
Lockery SR and Goodman MB. (1998). Tight-seal whole-cell patch clamping of Caenorhabditis elegans neurons. Meth. Enzymol. 293, 201-217.
Richmond JE and Jorgensen EM. (1999). One GABA and two acetylcholine receptors function at the C. elegans neuromuscular junction. Nat. Neurosci. 2, 791-797.
{kind=link}