2. Present address: Laboratory for Comprehensive Bioimaging, RIKEN Center for Biosystems Dynamics Research, Kobe, Japan
Proper positioning through attachment to the body is essential for tissue and organ function. We had previously reported the lethal mutant, ven (for ventral cord abnormal), exhibiting nerve cord displacement or detachment (Shioi et al., 2001). Herein, we report our identification of the ven-1 gene through genetic/physical mapping as well as transformation rescue of the Ven phenotype.
We cloned a genomic fragment that rescued the Ven phenotype (Figure 1A). All the mutated alleles were detected in the genomic region. From cDNA libraries, we isolated a 345 bp cDNA clone corresponding to the C53A.2 gene assigned on WormBase. RNA interference experiments using this cDNA sequence reproduced the Ven phenotype. The ven-1 transcript encoded a polypeptide of 86 amino acid residues, including the signal sequence at the N-terminus, suggesting that VEN-1 is a secreted protein (Figure 1B). VEN-1 appears to be conserved in Nematoda but not in other phyla.
To gain insight into ven-1 expression, we constructed ven-1p::GFP, in which the 472 bp upstream region of the ven-1 open reading frame was fused to green fluorescent protein (GFP) cDNA (Figure 2A). The GFP signal was first detected at the comma stage before ventral enclosure and remained until the 3-fold stage (Figure 2B). In the comma stage, it was expressed in hypodermal cells: Abpl/raapppp (H7), P1/2 L/R, P3/4 L/R, P5/6 L/R, P7/8 L/R, P9/10 L/R, and P11/12.
Next, we constructed ven-1::HA, in which the C-terminus of VEN-1 in the rescue fragment was tagged with an influenza hemagglutinin (HA) peptide. The ven-1::HA sequence rescued the Ven phenotype, and the HA epitope was detected in hypodermal cells (Figure 2B). The signal consisted of short oblique parallel lines arranged in four rows along the lateral body wall, corresponding to the position of muscle cells. The periodic nature of staining suggests that it may correspond to structures such as M lines and/or dense bodies (Moerman and Williams, 2006).
To examine the possibility of VEN-1 being a secreted protein, we tested whether forced expression of the ven-1 transcript under the control of heterologous promoters could rescue the Ven phenotype in tissues. The forced expression of the transcript in body wall muscle tissue rescued the Ven phenotype as efficiently as the genomic fragment did (Figure 2C). Its forced expression in neuronal and intestinal tissues also resulted in partial rescue of the phenotype. These results support the hypothesis that VEN-1 is a secreted protein.
We also examined the temporal requirement of ven-1 activity by expressing the transcript using a heat shock promoter. Its expression after egg laying completely rescued the Ven phenotype (Figure 2D). However, the recovery ratio gradually decreased after the comma stage, and expression of the transcript after the 1.5-fold stage failed to rescue the Ven phenotype. Therefore, ven-1 expression is required during the embryonic stages when nerves and muscles differentiate and attach to the body wall.
The ven-1 mutants also showed muscle attachment defects (Mua) (Shioi et al., 2001). Therefore, we examined the localization of attachment components of body wall muscles in ven-1 mutants using monoclonal antibodies against intermediate filaments (MH4), perlecan (MH3), and myotactin (MH46) (Labouesse, 2006; Moerman and Williams, 2006). Staining with the antibodies was detected in the displaced muscles (Figure 3), suggesting that ven-1 mutants have major defects in the hypodermis.
Previous studies have reported that a mutation in the gene encoding myosin-4 (unc-54) suppresses the Mua phenotype of mua-1 mutants (Plenefisch et al., 2000). However, we found that the ven-1(nc25); unc-54(n190) double mutant had a stronger phenotype than each single mutant, with its penetrance of the Ven phenotype being higher than that of the ven-1 mutant and its growth being much slower than that of the unc-54 mutant. These results indicate that the function of VEN-1 is different from that of MUA-1.
Figures
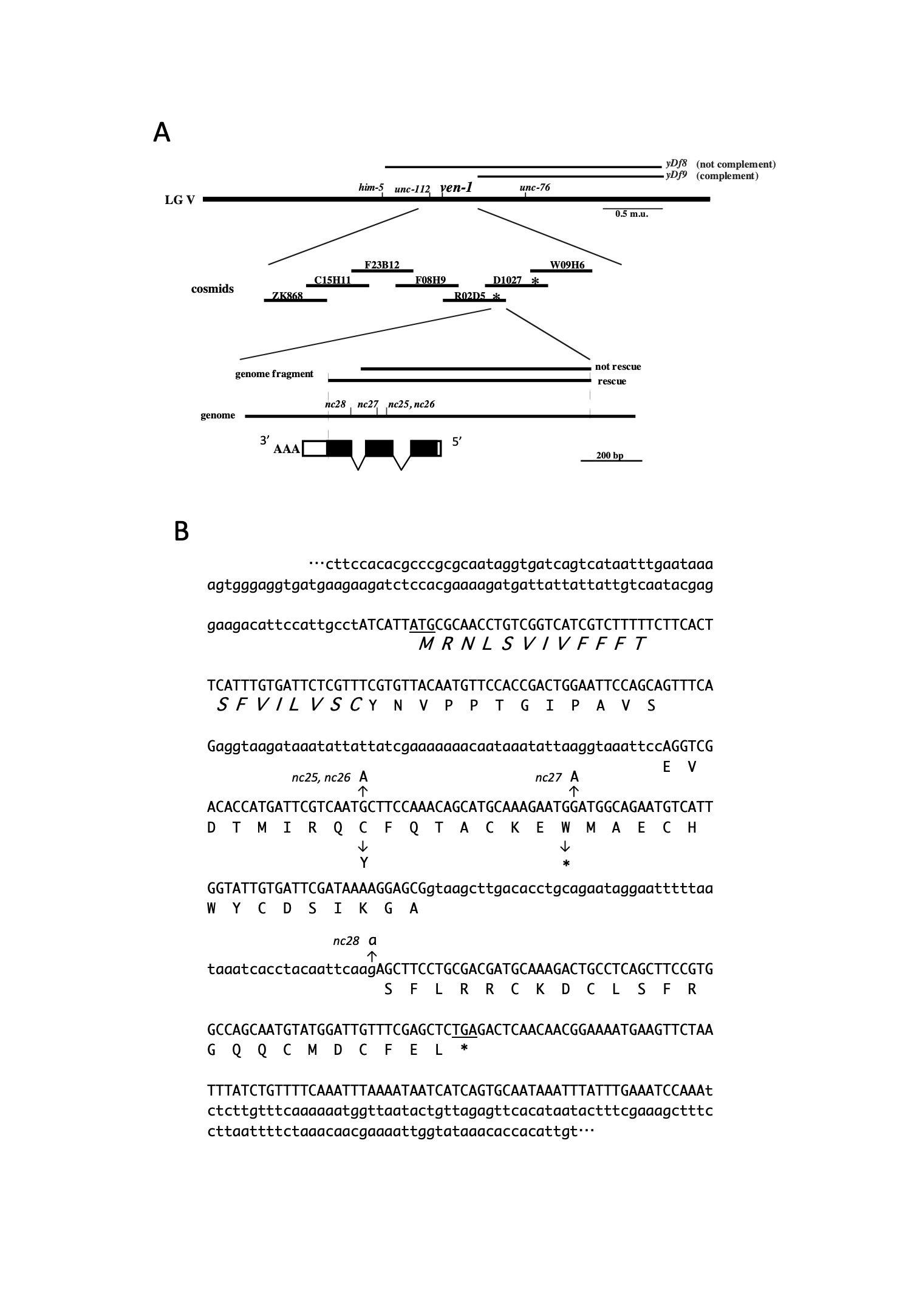
(A) Schematic representation of the genetic and physical maps of ven-1. All the ven-1 alleles were genetically mapped between unc-112 and unc-76. ven-1 was complemented by the deficiency yDf9, but not by yDf8. Two cosmids, R02D5 and D1027 (asterisk), and a 900 bp genomic fragment derived from R02D5 rescued the ven-1 defects. The ven-1 cDNA, composed of three exons, is shown at the bottom. Box: exons; black box: coding region of the ven-1 open reading frame (ORF).
(B) Sequence of the ven-1 transcript and ORF. The ven-1 transcript is 345 nt long and contains an ORF of 258 bp. The putative signal sequence is shown in italic font. nc25 and nc26 are missense mutations, whereas nc27 is a nonsense mutation. nc28 is a point mutation in the splicing acceptor on the second intron.
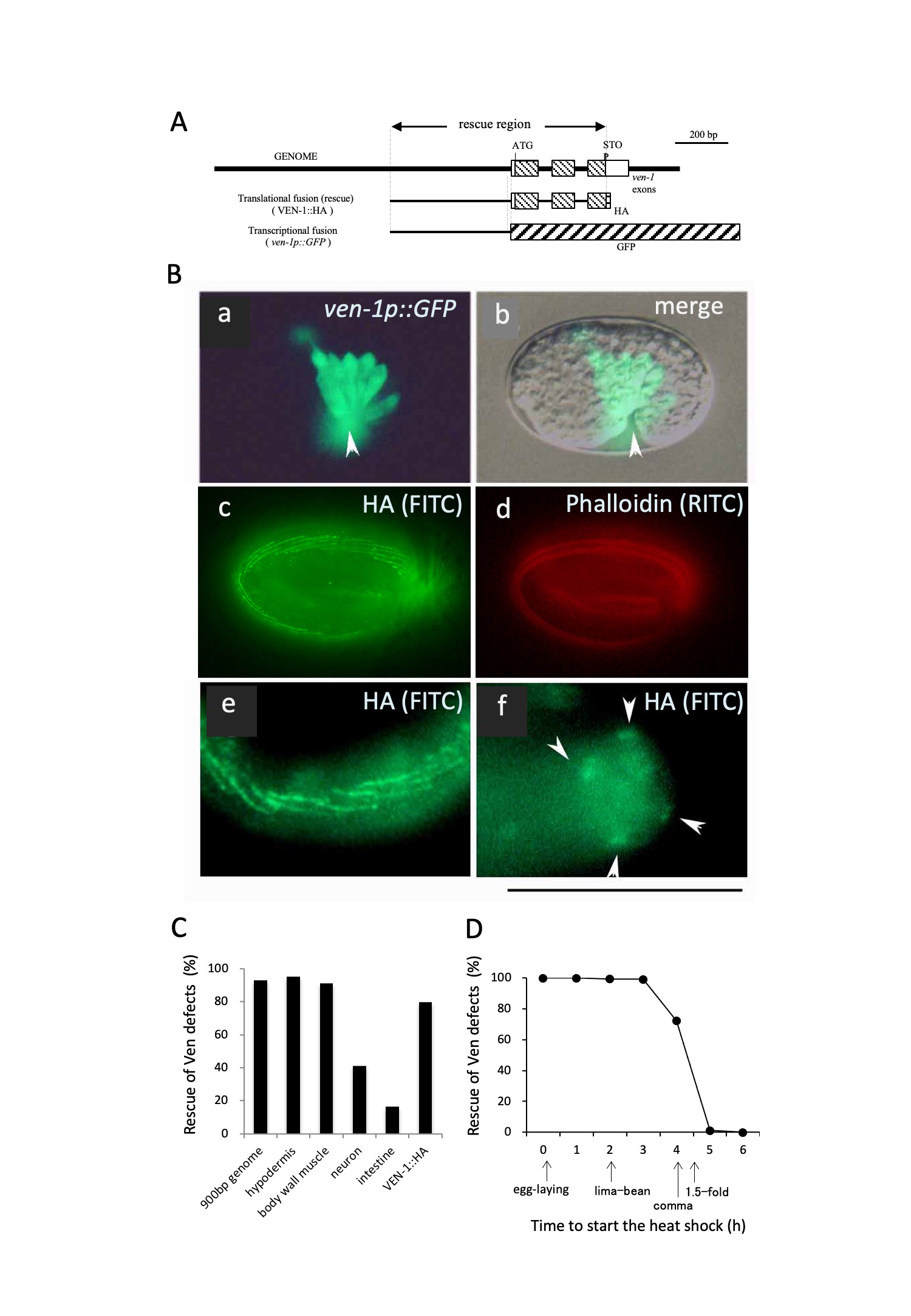
(A) Schematic illustration of the ven-1::HA and ven-1p::GFP constructs
(B) Expression pattern of ven-1. (a, b) The GFP signal in worms carrying ven-1p::GFP was expressed in ventral hypodermal cells from the comma stage. The arrowhead indicates the ventral side. (c, d, e, f) Worms expressing VEN-1::HA were doubly stained with the HA antibody (c, e, f) and phalloidin (d). Scale bar: 100 µm.
(C) Rescue experiments with ven-1 cDNA expressed in ven-1(nc25) under the control of heterologous tissue-specific promoters: epidermis (dpy-7 promoter, 94.7%, n = 227), body wall muscles (unc-54 promoter, 91.1%, n = 158), neurons (jkk-1 promoter, 40.9%, n = 227), and intestine (elt-2 promoter, 16.3%, n = 469). VEN-1::HA (79.6%, n = 141) and the 900 bp genomic fragment (93.0%, n = 127) corresponded to the rescue region in (A).
(D) Rescue experiments with ven-1 cDNA expressed in ven-1(nc25) under the control of a heat shock promoter. Eggs at 0–6 h after being laid were grown at 30°C until hatching: 0 h: 100.0%, n = 94; 1 h: 100.0%, n = 240; 2 h: 99.4%, n = 190; 3 h: 99.2%, n = 274; 4 h: 72.7%, n = 191; 5 h: 1.1%, n = 286; 6 h: 0.1%, n = 152.
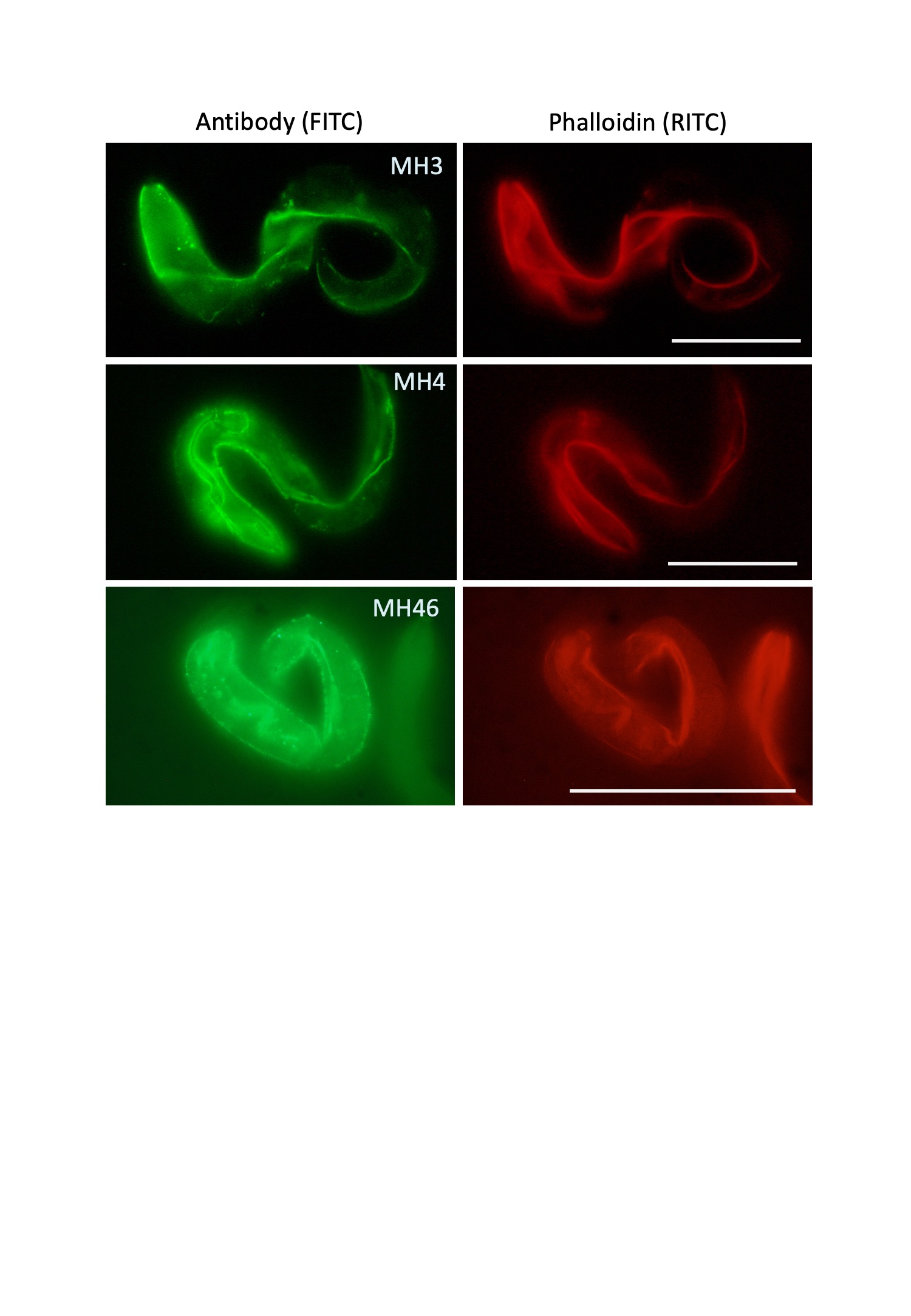
(Left) Antibody staining. MH4: intermediate filament; MH3: perlecan; MH46: myotactin. Immunofluorescence signals were detected on body wall muscles. (Right) Phalloidin staining. Scale bars: 100 µm.
References
Shioi, G., M. Shoji, M. Nakamura, T. Ishihara, I. Katsura, H. Fujisawa, and S. Takagi (2001). Mutations affecting nerve attachment of Caenorhabditis elegans. Genetics 157, 1611-1622. 
Moerman, D. G. and Williams, B. D. (2006). Sarcomere assembly in C. elegans muscle, WormBook, ed. The C. elegans Research Community, WormBook
Labouesse, M. (2006). Epithelial junction and attachments, WormBook, ed. The C. elegans Research Community, WormBook
Plenefisch, J. D., X. Zhu and E. M. Hedgecock (2000). Fragile skeletal muscle attachments in dystrophic mutants of Caenorhabditis elegans: isolation and characterization of the mua genes. Development 127, 1197-1207.
Articles submitted to the Worm Breeder's Gazette should not be cited in bibliographies. Material contained here should be treated as personal communication and cited as such only with the consent of the author.
Leave a Reply
You must be logged in to post a comment.