Worm Breeder's Gazette 9(3): 85
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Recently, we have been determining the time microfilaments (MFs) are
needed to generate and maintain proper asymmetry during the first cell
cycle of Caenorhabditis We can disrupt the
MF system using the MF inhibitor cytochalasin D (CD). As an indication
of asymmetry, we have been observing the behaviour of the zygote
during pseudocleavage, pronuclear migration, segregation of germ-cell-
specific granules (P granules), and first cell cleavage, all of which
clearly show some indication of asymmetry. As has already been
reported, when embryos are made permeable to CD early in the first
cell cycle all manifestations of asymmetry are destroyed:
pseudocleavage disappears, the egg and sperm yronuclei meet in the
center of the embryo, P granules fail to segregate posteriorly and the
first mitotic spindle sets up and remains symmetrically placed
throughout karyokinesis. If one-cell embryos are treated with CD after
the pronuclei have met and P granules have been segragated, P granules
remain posterior but the mitotic spindle sets up and remains symmetric.
This suggests that CD affects those events which have not yet taken
place, but does not reverse the asymmetries already generated. It also
suggests that MFs are not required to hold P granules in the posterior
of the embryo once they have been segregated there. They may be
anchored to non-MF components or may not be free to diffuse due to the
nature of the cytoplasm.
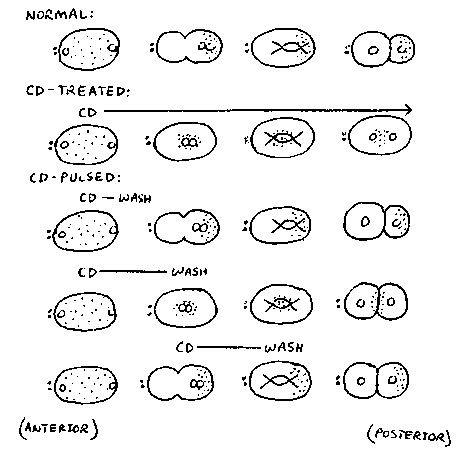
Zygotes can be pulsed with CD by introducing drug and then washing
the drug out of the zygotes at specific times. Control experiments
show that CD disrupts MFs within one minute of drug treatment. The
minimum time zygotes were treated with drug during experiments was two
minutes. Based on three types of CD pulse experiments (see
diagram), it appears that MFs are not required prior to pseudocleavage
and pronuclear migration but are required from pronuclear meeting
until cleavage to generate proper asymmetry. l)If the embryo is
treated very early, during pronuclear formation or early
pseudocleavage, and the drug is washed out two minutes later, the
embryo will resume pseudocleaving and behave like a normal embryo with
respect to all observable aspects of asymmetry. 2)If CD is added to
the embryo prior to pronuclear migration and washed out shortly after
the pronuclei reach the center of the embryo, the embryo is able to
recover enough to form a cleavage furrow and carry out cytokinesis.
However, the proper asymmetry of the cleavage is lost. Most often the
embryo divides into two equal-sized daughters, although a small
percentage of the time the furrow is formed to give either a slightly
larger AB-cell or a slightly larger Pcell. The P granules are not able
to recover from the treatment and remain localized in the center of
the embryo as they do in a continuously cytochalasin-treated embryo. 3)
Preliminary results show that embryos treated shortly after pronuclear
migration and P granule segregation and then washed several minutes
later also undergo symmetric cleavage. However, in these embryos, P
granules are segregated to the posterior cell. We conclude from these
experiments that proper MF structure is needed at a critical time,
during late pseudocleavage and pronuclear migration, in order for
pronuclei to meet properly and P granules to segregate properly. After
this time, microfilaments are needed in order to generate proper
asymmetry in the first cell division itself but are dispensible with
respect to events which have already occurred (P granule localization).
Further experiments will be done with various cytochalasin-pulsed
embryos to examine how pulses affect the developmental potential of
daughter cells.
{Figure 1}