Worm Breeder's Gazette 9(3): 70
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
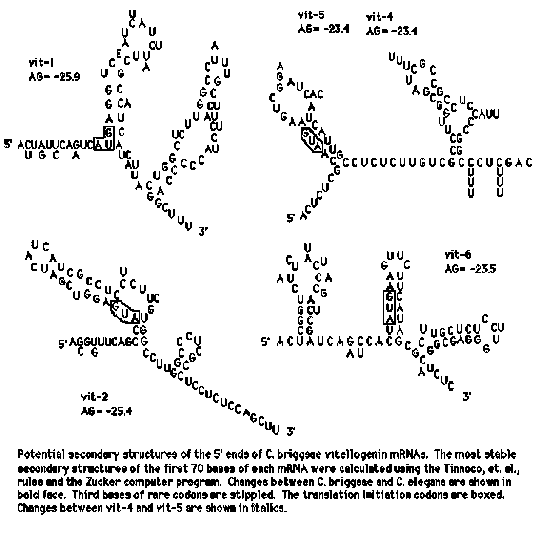
We asked the computer to find the most stable potential secondary structures that the 5' ends of the vitellogenin mRNA's could fold into. It found quite stable structures ( G congruent to -24 Kcal/mol) in the first 70 bases of vit-2, vit-5 and vit-6. The same region of vit-1, which we know is a pseudogene, could not be folded into a structure of equivalent stability. In each case, the AUG at which translation initiates is involved in a hydrogen bonded structure. Do these structures actually form and, if so, what are they for? We have three lines of evidence that they have been selected for and therefore presumably perform some function. (1.) We have sequenced the equivalent regions of the C. briggsae genes and found that the proposed stem-forming regions have been highly conserved. Most base pair changes between C. elegans and C. briggsae are in unpaired regions; while the unpaired regions have diverged by about 20%, approximately the same as the rest of the coding regions we have sequenced, the sequences proposed to be involved in stem formation have diverged by only 2%. Furthermore, the few changes in the stems that have occurred would not result in alteration of stem stability. (2.) Although vit-1 is a pseudogene in C. elegans, it is apparently an expressed gene in C. briggsae and, in the latter, a stable stem-loop structure is predicted. There are far more base changes between the two species in vit-1 than in the other gene pairs, but even in vit-1 a preponderance of the changes are in unpaired regions. This may reflect the fact that vit-1 became a pseudogene in C. elegans relatively recently. (3.) We have noted before (Spieth, et. al., 1985, Nucleic Acids Res. 13, 7129) that the 5' most 70 bases of all of the vit genes contain a much higher frequency of rare codons than do the rest of the coding regions. Although we originally had no satisfactory explanation for this finding, we believe it can be understood in the context of the step-loop structures. It turns out that the great majority of third- position bases which result in the presence of a rare codon are predicted to be involved in stem formation. In most cases, if these bases were changed to ones resulting in common codons, the stems would be significantly destabilized. Thus, it appears that selective pressure to maintain the ability to form the stem-loop structures is greater than the pressure to utilize certain codons. We have two hypotheses for what functions the stems might perform: First, they might be responsible for mRNA stabilization. Although we don't know whether the nematode vitellogenin mRNA's are especially stable, it has been shown that their close relatives, the Xenopus vitellogenin mRNA's are quite stable as long as estrogen is present. Interestingly, we have found that structures very similar to those shown in the Figure can be drawn for the 5' ends of all of the vertebrate vitellogenin mRNA's. The second possibility is that the proposed secondary structures are involved in regulation of vitellogenin translation. It may be that under some conditions, starvation for instance, that it makes sense to cease banking such a large quantity of amino acids for the next generation, but it doesn't make sense to destroy the large amount of vitellogenin RNA already synthesized. Thus, it may be that the system is designed to modulate vitellogenin translation as a response to environmental conditions, perhaps by formation of secondary structures close enough to the 5' end to prevent ribosome loading. Experiments to test these fantasies are in progress. [See Figure 1]