Worm Breeder's Gazette 9(2): 57
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
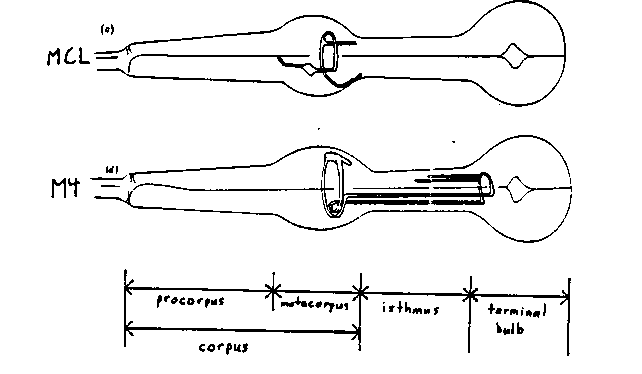
The pharynx has its own nervous system consisting of 20 neurons. There are eight unpaired neurons and six bilaterally symmetric pairs, making a total of fourteen classes (Albertson and Thomson, Phil. Trans. Roy. Soc. London , 275: 299-325, 1976). We are using a laser (see the note by Avery, Herman, and Horvitz in this issue) to ablate sets of pharyngeal neurons in the hope of learning how the pharyngeal nervous system works. We looked for obvious changes in the rate of pumping of worms on plates with bacteria, for changes in the rate of growth, and for bloating caused by defects in egg-laying. ( The rate of egg-laying is regulated by the presence of bacteria, which might possibly be detected in the pharynx.) Unless otherwise stated, all ablation experiments described below were replicated in at least five animals. We also used Nomarski microscopy to check one or two animals from each experiment for changes in the motions of specific pharyngeal muscles. Most of the ablations were done in N2 L1 hermaphrodite larvae within a few hours of hatching. We began with single class ablations, in which only one of the fourteen classes was ablated in a single worm. Ablation of two classes of neurons, MC and M4, had striking effects, described below. However, by the tests described above we saw no effect of ablation of any one of the remaining twelve classes: I1, I2, I3, I4, I5, I6, MI, M1, M2, M3, M5, and NSM. This lack of effect is partially due to redundancy: when eleven of the twelve classes (all but I1) were ablated in each of two animals, pumping was slowed and development retarded, as described below for MC-ablated worms. However, these animals did pump, and in fact grew to adulthood and made progeny. Even when all 20 pharyngeal neurons were ablated, the worms still pumped. Moreover, as in intact animals, the corpus and terminal bulb ( see figure for the names of parts of the pharynx) contracted at the same time in animals lacking a pharyngeal nervous system. We can explain this result by proposing that all the pharyngeal muscles are electrically coupled and capable of autonomous contraction, like a vertebrate heart. The two MC neurons MCL and MCR form a bilaterally symmetric pair ( see figure). In young worms their nuclei are in the ventral metacorpus, anterior to the NSMs and dorsomedial to the I2s (J. Sulston, personal communication) The MCs innervate the marginal cells, whose function is unknown. When both MCs were ablated, the worms pumped slowly and development was slightly retarded: at 20 C the worms took three days to reach adulthood from hatching, instead of the usual two. The worms looked slightly starved (small size, lack of brown color in the intestine) as larvae and young adults, and very starved while they were producing eggs. By the crude criterion of failure to bloat, all laid eggs normally. Sulston and Horvitz had also ablated MC and seen similar effects (personal communication). M4 is an unpaired motoneuron, the most posterior of the three in the dorsal metacorpus. M4 innervates the posterior half of the isthmus ( see figure). The isthmus regulates the passage to the terminal bulb of bacteria accumulated in the corpus, propagating peristaltic waves that carry pellets of bacteria to the front of the grinder. When M4 was ablated, the posterior half of the isthmus remained closed. The worms still pumped, but the corpus and anterior isthmus quickly became stuffed with bacteria, while the grinder operated on nothing. As in intact worms, the terminal bulb and the corpus contracted synchronously. The anterior isthmus opened normally during corpus contractions. Animals lacking M4 arrested development as L1 larvae. One animal continued to be active and apparently healthy for nine days, but most worms lacking M4 crawled off their plates within a few days. M4 is the anterior daughter of MSpaaaaa, whose posterior daughter, MSpaaaaap, dies in N2 (Sulston, Schierenberg, White, and Thomson, Dev. Biol. 100: 64-119, 1983). The mutation ced-3(n717) prevents this death. In about a third of ced-3 L1s, we saw two morphologically similar nuclei clearly visible where M4 is in N2, one anterior to the other. (MSpaaaaap may have been somewhere else in the remaining two thirds of the worms, or the ced-3 mutation might be leaky.) When both of these nuclei were ablated, the effect was similar to that of ablating M4 in N2. When the posterior of the two was ablated and the anterior left intact, all the animals grew and pumped normally. However, when the anterior nucleus was ablated, leaving only the posterior one, most of the animals grew very slowly, and some did not become adults or produce progeny. Our interpretation of these results is that, when the two cells are near the normal M4 position, the anterior one is usually M4, and the posterior MSpaaaaap. MSpaaaaap is not functionally identical to M4. However, most M4 MSpaaaaapt ced-3 worms did better than either M4 MSpaaaaap ced-3 or M4 N2, so we suspect that MSpaaaaap can sometimes partly replace M4.