Worm Breeder's Gazette 9(2): 40
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
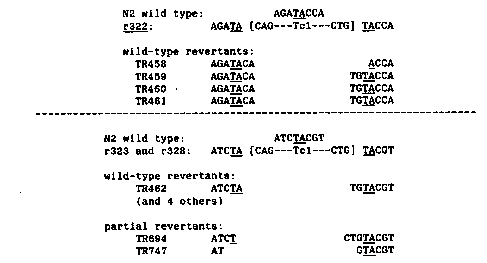
Many Tc1-induced mutations are unstable in the germ line and revert
at high frequencies. We have sequenced nine wild-type and two partial
revertants of three Tc1-induced unc-54 mutants. Southern blots showed
that these revertants contained precise or nearly precise excisions of
Tc1, but we sequenced them to determine their exact structures. The
results are summarized below. Revertants of r328 and r323 are grouped
together because these insertions are in the exact same site. The
putative target site duplications are underlined and the limits of Tc1
are indicated with
brackets:
[See Figure 1]
Each revertant results from imprecise excision of Tc1, and each
sequence can be generated by simple deletion of Tc1 or adjacent target
site material.
The Tc1 insertions used in this study are in exons. This may limit
our ability to detect excisions because only certain imprecise
excisions will yield revertant phenotypes. For example the four wild-
type revertants of r322 have either three or six base insertions
relative to the wild-type gene corresponding to one or two added amino
acids. Thus, the translational reading frame is maintained at this
site. The r322 insertion is in a region of the myosin gene encoding
the rod portion of the protein and the additional amino acids in this
region cause no major loss of function.
Five wild-type revertants of r323 and r328 are identical and contain
four base insertions relative to the wild-type gene. We believe that
the following explanation (described for TR462 in the previous
newsletter) accounts for their revertant phenotype: The target TA
dinucleotide is one base away from the 5' splice site of rV5#3. We
suggest that these revertant alleles contain a new splice site that is
displaced four bases 5' of the normal splice site. Thus the four base
insertion is spliced out of the mRNA. The sequence of the putative
new splice site fits well with the C. elegans splice site consensus
compiled by Tom
Blumenthal:
[See Figure 2]
The putative new 5' splice site is indicated by the vertical line (1)
The normal splice site is indicated by the apostrophe ('). Splicing
at the new site would generate a protein whose amino acid sequence is
identical to the wild-type protein. These revertants contain normal
or near normal levels of unc-54 protein.
The partial revertant TR694 also has a four base insertion. As
above, a putative new 5' splice site is
generated:
[See Figure 2]
In this case, however a single amino acid substitution (tyrosine to
serine) is generated. Protein gels show that unc-54 protein levels
are approximately wild type in TR694 so we believe that the partial
revertant phenotype is due to the amino acid substitution and not a
splicing defect. The r323 insertion is in the head region of the
protein close to the ATP-binding site.
The partial revertant TR747 differs from wild type by a single base
substitution. This alteration causes an amino acid substitution and
also creates a new potential 5' splice displaced by four base pairs.
[See Figure 2]
However since the revertant allele does not contain any insertion or
deletion relative to the wild-type gene use of the new splice site
would result in a translational frameshift at this site. The level of
unc-54 protein in TR747 is reduced about 5-8 fold relative to wild
type so we suspect that the new splice site is preferentially utilized.
An alternative possibility is that the amino acid substitution
destabilizes the protein. Experiments are in progress to analyze the
pattern of splicing in these various revertant strains.
It is important to consider the frequency of the reversion events
discussed above. Wild-type revertants of r323 are 10- to 100-fold
more frequent than either the partial revertants of r323 or the wild-
type revertants of r322. The relative frequencies of these reversions
likely reflect the ability of each insertion site to tolerate
imprecise excision while restoring gene function. The most frequent
type of Tc1 excision that we detect accounting for >99% of r323 germ
line reversion events leaves a specific four base pair insertion (TATG)
at the point of excision. We detect these events because the r323
insertion site is especially tolerant of imprecise excision.
Excisions of this type would not be detected for many Tc1 insertions.
Such sequences may represent the most frequent type of Tc1 excision.
The TATG insertion is the same as that observed by Ruan & Emmons for a
somatic excision event of a natural Bergerac Tc1 element.
