Worm Breeder's Gazette 9(1): 66
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
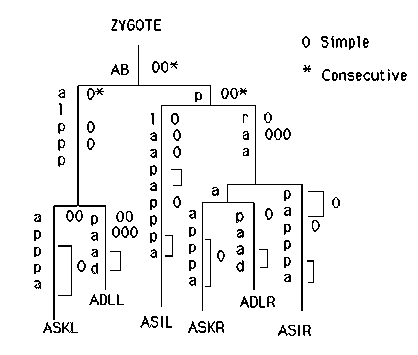
Recessive ncl-1 mutations, e1865 and e1942, cause enlargement of the nucleoli of all cells. The phenotype can be scored by Nomarski microscopy and homozygous mutants are otherwise normal and fertile. If cell autonomous, ncl-1 should be a general, non-destructive marker for genetic mosaicism. sDp3 is a free duplication covering the left arm of LGIII that was generated by R. Rosenbluth, C. Cuddeford and D. Baillie. Recently, C. Kenyon has shown that sDp3 is mitotically unstable and has used it to study mosaics. As sDp3 covers ncl-1 (and dpy-1), we constructed a strain of genotype sDp3; , 65) to examine the autonomy and perdurability of the ncl-1 phene. We scored six amphid neurons in young, non-dpy adults (Figure 1). Our choice was based on the ease of scoring these cells and their wide separations in lineal origin. From roughly 240 animals, we identified 30 in which one or more of these neurons were ncl-. Scoring additional cells and using the published embryonic cell lineage, we could interpret most as simple mosaics resulting from duplication loss by a single precursor (Figure 1). As noted for other duplications, the probability of loss appears to be roughly constant throughout the lineage. In a few cases (brackets in Figure 1), loss could only be narrowed to a cell or its mother because the sister had subsequently fused with ncl+ cells. In syncytia formed by the fusion of presumptive ncl- and ncl+ cells, all nuclei had the ncl+ phenotype. No exceptions to the published lineage were observed. As expected, some mosaic animals contained additional clones of ncl- cells arising by independent loss of sDp3 at two or three consecutive divisions. This frequency of consecutive mosaics appears too high to be explained by multiple, independent events. A simple mechanism for duplication loss is for replication or segregation to fail at one cell division, yielding a daughter with one or two duplications and a daughter with no duplication. If replication and segregation recover for the next cell division, a simple mosaic should result. If these functions do not recover until further division has occured, a consecutive mosaic might be produced. Conceivably, some animals interpreted as simple mosaics actually represent consecutive mosaics in which a ncl+ subclone was overlooked. Newly hatched dpy-1(e1) animals are normal but they lengthen less than wild-type during larval growth and, as adults, are slightly irregular in girth and have abnormal alae. Duplication loss in AB ( four cases) results in normal length animals with irregular anterior alae and a ventral bend in the extreme tail. Duplication loss in ABa ( two cases) has no effect and loss in ABp (three cases) results only in the ventral tail bend. Three animals picked for having a ventral tail bend proved to be AB or ABp mosaics. Duplication loss in P1 (one case) produced a fully normal animal except for a dorsal tail bend. In summary, expression of dpy-1+ in either AB or P1 is sufficient to produce an animal of normal length. This suggests that the site of dpy-1 expression is the hypodermis and, in particular, the hyp7 syncytium comprising both AB and P1 descendants. We have speculated that the normal ncl-1 product may be a repressor of rRNA transcription. Interestingly, the ncl- neurons in mosaics are much larger than their ncl+ neighbors. (The ncl-1 mutants appear to be somewhat larger than wild-type.) The increase in cell size may reflect an increase in the number of functional ribosomes and general protein synthesis. I wish to thank R. Rosenbluth for kindly providing sDp3(III;f), R. Herman and C. Kenyon for their generous encouragement. CB3461 (ncl-1( e1865)) is available from the CGC.