Worm Breeder's Gazette 9(1): 12
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
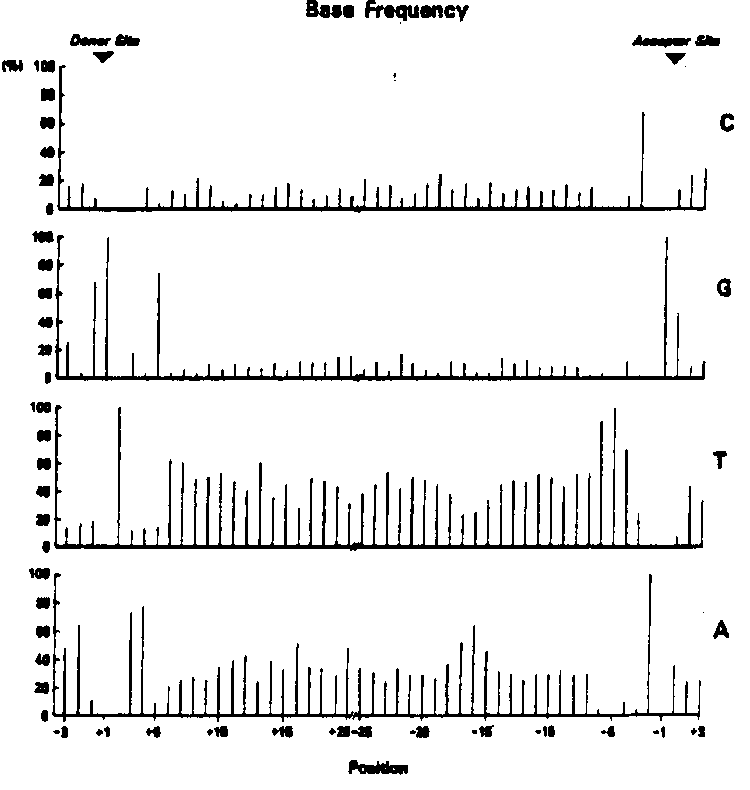
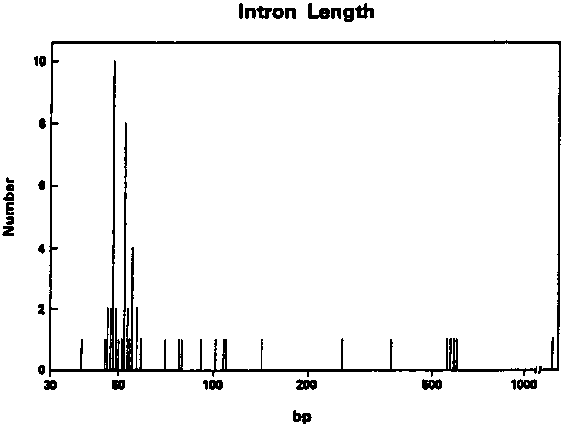
With the cooperation of several C. elegans gene sequencers, I have collected data on 53 worm introns. This is a brief report on some of the similarities and differences between introns of C. elegans and those of other metazoans. 1. C. elegans introns are unusually short (Fig. 1). Three quarters are between 45 and 59 bases long, and the majority of the very short introns are either 48 or 52 bases in length. Interestingly, experiments with introns from higher eukaryotes have suggested that introns shorter than about 80 bases cannot be spliced. This suggests the mechanism for splicing C. elegans introns may be somewhat different from that used for splicing introns from higher eukaryotes. 2. C. elegans introns are very rich in A+T. They average 74% A+T, and only two of the 53 introns analyzed are under 67% A+T. 3. The C. elegans border sequences are similar to those of other metazoans, but there are significant differences (Fig. 2). The donor sequence is nearly identical to the general eukaryotic consensus, while the acceptor sequence is more atypical: there is a strong preference for T's at positions minus four through minus six. All other positions in the intron have a consensus of A/T, including the eleven positions near the 3' splice junction where the consensus is C/T in other eukaryotic introns. [See Figure 1] [See Figure 2] 4. The presence of an internal consensus sequence, PyTPuAPy at which lariat formation occurs in other eukaryotic introns from yeast to man is not a characteristic of C. elegans introns. Nearly half of the short introns lack this sequence. Nor is there any obvious different consensus sequence to replace it. However there is a preference for A at positions minus 15 through minus 17 in the intron ( Fig. 3). Since lariat formation always occurs at A residues, and since the lariat consensus is only weakly adhered to in other systems, it may be that lariat formation occurs at the A's between minus 15 and minus 17. It should be noted however that these positions are closer to the 3' boundary than the lariat sites of other introns (minus 22 through minus 37). [See Figure 3] In conclusion, the overall similarity between the border consensus of C. elegans and those of other eukaryotes suggests that the mechanism of splicing nematode introns will turn out to be very similar to those being worked out in mammalian systems. On the other hand, the small size of C. elegans introns, their somewhat unusual 3' boundary sequence, and their lack of a canonical lariat consensus sequence suggest that there are some special aspects of splicing in nematodes which may justify an experimental attack on the problem. The fact that so many C. elegans introns are clustered around 50 bases in length suggests that this may be an optimal size for efficient splicing, and/or that C. elegans genes experience selective pressure resulting in intron length compression.