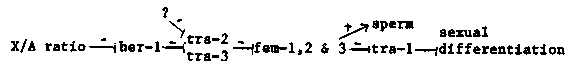
Worm Breeder's Gazette 8(3): 34
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
One of the unsolved problems in understanding how genes control sex determination in C. elegans is that of how spermatogenesis, a male function, occurs in the hermaphrodite, which is otherwise female. The current model for sex determination can only explain how male vs. female development occurs. As described in the last Newsletter, I have been working on several dominant tra-2 mutations that may bear on this problem. These mutations eliminate spermatogenesis in XX animals, to produce females instead of hermaphrodites, but do not generally show effects on XO males. In the XX animal, tra-2 is thought to act with tra-3 to prevent the function of the fem genes. In the XO animals, tra-2 appears to be turned off by the action of her-1, thereby allowing the fem genes to promote male development and spermatogenesis. Since the fem genes are also needed for spermatogenesis in the XX animal, there must be a way of allowing them to promote a brief period of spermatogenesis without interfering with female somatic development or eventual oogenesis. [See Figure 1] The tra-2(dom) mutations indicate that this transient fem expression is mediated via the modulation of tra-2 activity. That is, that although tra-2(+) is on in the XX animal, it is turned off transiently to allow the fems to promote spermatogenesis. In tra-2(dom) animals, tra-2 activity is apparently no longer modulated, i.e. it is constitutive, because tra-2(dom) XX animals are females. However, XO tra-2(dom) animals are usually normal males, therefore tra-2(dom) is still turned off by her-1 in the XO animal. In sum, in addition to the negative control of tra-2 activity by her-1 in the XO animal, tra- 2 is also modulated in the XX animal to allow hermaphrodite spermatogenesis to occur. It is possible that genes that specifically control this modulation exist (indicated by the ? in model). Two tra-2(dom) alleles, tra-2(e1940) and tra-2(e2020), have been characterized in greater detail. Both have been reverted to yield several intragenic recessive masculinizing mutations that fail to complement tra-2(e1095) (a putative null). This indicates that they are tra-2 alleles. Although both mutations appear to be constitutive, their penetrance differs: tra-2(e1940) and tra-2(e1940)/+ XX animals are sometimes slightly self-fertile hermaphrodites instead of females, while tra-2( e2020) and tra-2(e2020)/+ XX animals are always female. Further, tra-2 activity appears to differ in the two alleles; tra-2( e1940) activity is possibly lower than wildtype (hypomorphic) because: 1) tra-2(e1940)/(e1095) XX animals are self-fertile intersexes in contrast to tra-2(+/e1095) XX animals, which are essentially wildtype 2) the masculinizing effect of her-1(n695sd) is enhanced so that tra- 2(e1940)/+;her-1(n695)/+ XX animals are more intersexual than her-1( n695)/+ alone. By contrast, tra-2(e2020) does not appear to be a hypomorph because tra-2(e2020)/(e1095) XX animals are female, in both soma and germline. tra-2(e2020) activity may even be higher than wildtype (hypermorpic) 1) tra-2(e2020);her-1(n695) XX animals are female 2) the masculinizing effect of the tra-3(e1767) mutation is partially suppressed so that tra-2(e2020)(/+);tra-3(e1767) animals will propagate slowly as a male/female strain. This result is significant because it indicates that tra-2(e2020) can substitute for tra-3(+) function. Another feature of tra-2(e2020) is that although tra-2(e2020) XO animals are morphologically male, they begin to produce oocytes and yolk (as judged by Nomarski microscopy) after spermatogenesis as mature adults. This indicates that control by her-1 may have been somewhat affected in tra-2(e2020) XO animals. In addition to indicating that the tra-2(dom) mutations affect XX- specific control of tra-2 activity, these results show that the tra-2( dom) mutations affect somatic as well as germline tissues. The fact that in some circumstances these mutations appear to affect only the germline suggests that the soma and the germline may have different thresholds of sensitivity to tra-2 activity. Dominant extragenic suppressors of tra-2(e1940) and tra-2(e2020) have been sought in the hope of identifying genes that act to modulate tra-2 specifically in the XX animal, allowing spermatogenesis. No suppressors of tra-2(e2020) have been found, but tra-2(e1940) has yielded two so far. Both are unlinked to tra-2(e1940) but are as yet unmapped. These mutations produce a novel phenotype, the 'sperm whale' (Sw), which is a hermaphrodite whose gonad makes only sperm. One of them affects the soma as well as the germline so that tra-2( e1940);sw/+ XX animals are self-fertile intersexes. sw/+, sw/sw and tra-2(e1940);sw XX animals are sperm whales or hermaphrodites, with variable penetrance. Finally, a mutation that appears to have germline-specific effects has appeared by chance in a suppressor hunt. This mutation, e1959, which maps to the left arm of LGI, is recessive in XX animals, producing females, and has a semidominant feminizing effect on the germline of XO animals. e1959/+ XO animals produce oocytes in a male gonad after a period of spermatogenesis, and e1959 XO animals often make only oocytes in a male gonad. No yolk production is seen by Nomarski microscopy.