Worm Breeder's Gazette 8(3): 18
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
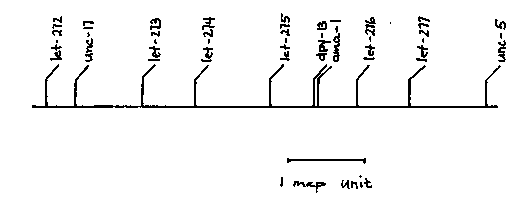
Sanford, Golomb and Riddle (J. Biol. Chem. 258:12804-12809, 1983) have identified a gene, ama-1 IV which appears to code for a subunit of RNA polymerase II. This gene is defined by the dominant mutation m118 and has been positioned near dpy-13 on LG IV. Strains carrying ama-1(m118) are resistant to alpha-amanitin and produce an altered RNA polymerase II enzyme. To continue the analysis of RNA polymerase in C. elegans we have (1) positioned ama-1 relative to dpy-13, (2) isolated lethal alleles of ama-1 and (3) identified several essential genes near ama-1. ama-1 was positioned relative to dpy-13 by testing Dpy recombinants from dpy-13(e184) + unc-5(e53)/+ 8)+ hermaphrodites for resistance to alpha-amanitin. The results obtained (2/154 Dpy's carried m118) place ama-1 approximately 0.03% to the right of dpy-13. The dpy-13(e184)ama-1(m118) double mutant constructed above was used in a screen to isolate lethal alleles of ama-1. Our strategy for isolating lethal ama-1 alleles was to first isolate lethal mutations on a chromosome carrying the dominant alpha-amanitin resistant allele m118 linked to dpy-13, and then screen each lethal strain for loss of resistance. This screen assumes that null alleles of ama-1 are lethal and that a second mutation in ama-1(m118) eliminating gene function would also eliminate resistance to alpha-amanitin in ama-1(m118mx)/+ heterozygotes. Briefly, homozygous dpy-13(e184)ama-1(m118) hermaphrodites were mutagenized (0.025M EMS) and then mated to N2 males. Strains carrying a lethal mutation on the dpy-13 me were identified by the absence of fertile, Dpy F2 progeny. A total of 35 lethal strains were isolated after screening the progeny of 3243 F1's. Each lethal strain (genotype: dpy- 13(e184)ama-1(m118)let-?(mx)/+++) was then tested in alpha-amanitin wells and six of the strains were found to be sensitive to alpha- amanitin. All six mutations define one complementation group and are presumably second site mutations in ama-1(m118). Two-factor recombination mapping of one of these mutations placed it 0.05% to the right of dpy-13. These six ama-1 alleles arrest development of the dpy-13(e184)ama-1(m118mx) homozygotes at various stages. Two are apparent embryonic lethals, two are late larval lethals, one is a temperature-sensitive sterile and the remaining mutation is not lethal but grows slowly and produces very few progeny. We are planning to do reversion experiments with these lethal ama-1 alleles in an attempt to identify genes coding for other RNA polymerase II subunits, or other genes affecting transcription efficiency. The remaining 29 lethal mutations that we have isolated define other essential genes in this region of LGIV. Left-right positioning and complementation data for 15 of these mutations have identified 6 essential genes in addition to ama-1. The positions of these genes ( based on two-factor mapping of one representative allele) are shown on the accompanying genetic map. We have recently isolated a set of gamma-ray-induced lethal mutations linked to dpy-13(e184)ama-1(m118), one of which affects ama- 1 as determined by loss of resistance to alpha-amanitin and failure to complement one of the ama-1 lethal alleles. It also fails to complement let-276 which is located approximately 0.5-map unit to the right of ama-1.[See Figure 1]