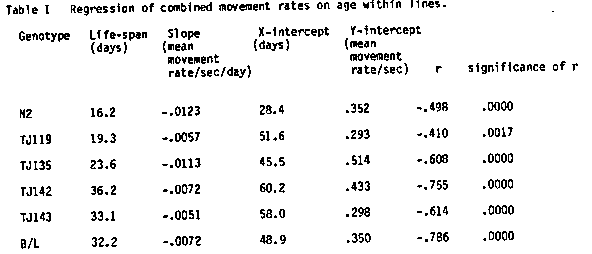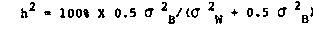
Worm Breeder's Gazette 8(3): 14
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
We followed movement rates in four recombinant inbred (RI) strains ( TJ119, TJ135, TJ142, TJ143) and the parental strains (N2 and Bergerac/B0) as a function of age to determine whether movement behavior can be used as a quantitative predictor of life-span. In preliminary experiments to determine the best methods for studying movements we counted forward waves, backward waves, and omega turns ( head to tail directional changes), and determined the reproducibility of four possible methods of assay. Three investigators independently scored individual movement of twenty 3-day old N2 hermaphrodites in four different media: 1) Undisturbed in liquid culture of S buffer plus E. coli (10+E9/ml) 2) Transferred from 1 to S buffer. 3) Transferred from 1 to plates prepared according to the Bolanowski et al. [Mech. Age. Dev., 15:279] method 4) Transferred from 1 to unspotted NGM plates. In all cases behavior was assayed at room temperature for 1 minute. Analysis of variance showed highly significant differences between media, but no significant differences between investigator for media 1, 3, and 4. Unspotted NGM plates were chosen for convenience for the study of age-dependent behavior using test populations of ten worms each from synchronous lays of the six genotypes. Our assays are of spontaneous, unstimulated rates of movement. We established aging cultures of the six strains. Each worm was maintained individually in microtitre wells under our standard conditions [P.N.A.S. 79, 6603] in 0.6 ml of media. Movement was observed for 60 seconds. Statistical analysis of a combined movement rate, the sumrate, (forward rate + backward rate + omega rate) yielded the following information: The mean movement rates showed significant negative correlations with chronological age for all genotypes, P<.002, (See Table I). Across the six RI lines, Pearson correlations between life-span and the slope of the regression, the X-intercept, or the Y-intercept were determined. Only the correlation with the X-intercept was statistically significant (see Table II) although slope was also highly correlated with life-span. Thus the X-intercept of the regression line of movement rate with age could be used as a predictor of life-span for these genotypes. These correlations indicate that the genotype of a strain has phenotypic effects that are expressed both in the life-span and in the ability to move as an individual worm ages. This is important because there are no accurate predictors of life-span in any organism and there are theoretical arguments for the lack of any predictors. Even within an RI line (genetically identical and therefore no genetic but only environmental influences) we observed the regression of movement rate with age for individual worms and the projected X-intercept to be positively correlated with life- span and thus potentially useful as a predictor. Heritability estimates of movement at each age indicated a high heritability during the reproductive period (40%) which diminished to near zero by day 11. Heritability (h ) was calculated using the [See Figure 1] as described by Johnson and Wood. This is an unexpected finding and indicates that the genetic influences on behavior become less dramatic with age; perhaps because the environmental components have more time to influence individual behavior. These results suggest that the senescent program may be played out by. mechanisms that have common effects on life-span as well as on behavior or conversely that behavior and life-span are causally connected. [See Figures 2-3]