Worm Breeder's Gazette 8(3): 12
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
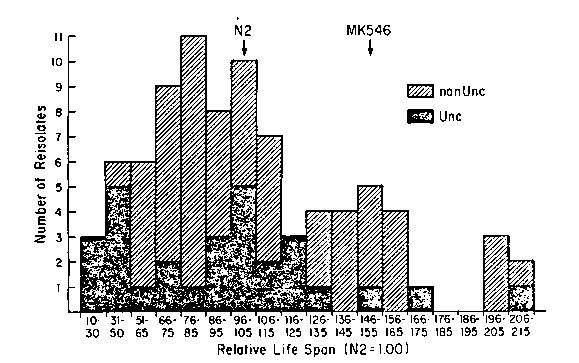
Klass isolated and completed preliminary characterization of several long-lived mutants isolated in DH26 by a brute force screening procedure (Mech. Age. Dev. 22:279). Mike ascribed the extended life-span to the secondary effects of either dauer formation or a decreased rate of food ingestion. Those mutants with decreased food ingestion also looked uncoordinated. We have asked which Unc locus(i) was involved in this increase in life-span (Age) and begun an attempt to map the Age locus. MK546, one of the longest-lived isolates, was crossed to N2 males and the F1 were allowed to self for four generations yielding stocks for analysis. We chose this procedure because pilot experiments suggested that few F2 animals were longer-lived; inbreeding to the F5 produces progeny which are 94% homozygous. Our data shows that the Unc phenotype is a single nuclear gene and maps to linkage group IV near dpy-13. It is allelic with unc-31 (e169). Complementation tests between e169 and Unc's of MR546, MR542, and MR31 (all isolated by Klass) showed that the Unc's were all allelic. Tests to determine allelism of the Age loci are currently underway. We tested the F5 for life-span and found that the Age phenotype segregates as two unlinked nuclear genes (X =0.140; p=0.7); this assumes that life-spans 1.26 times those of wild-type and above are Age and all others are non-Age and that there is no selection against the Age phenotype (see Figure). MR546, in addition to being Unc and Age, developed slowly, had small brood sizes and high frequencies of bags. We tested for correlations between Age and these other phenotypes in the F5. We found that Age is not linked to Unc (X2 =4. 12; p=0.25, see Figure). Age and small brood size do not cosegregate, nor do Age and high incidence of bags, nor do Age and slow development rate, nor do Age and b26. Not surprising was the correlation between Unc and both higher incidence of bags and slow development rate. Food uptake was monitored by ingestion of [35S] labeled 0P50 and compared with N2 controls and zero time samples; pharyngeal pump rate was also monitored. We found no difference between food uptake in MR546 and the other Mk strains as compared with wild-type or DH26. Comparisons of Age reisolates and wild-type reisolates also failed to detect differences in rate of food uptake. Extensive repeats of these observations lead us to accept their validity. We are attempting to localize the two genes responsible for the Age phenotype to their respective linkage groups (not an easy task). Age strains containing Unc's shown to have wild-type life-span representing each linkage group have been constructed by four rounds of backcrossing F2 Unc hermaphrodites with a male fertile, Age reisolate strain (TJ401). This eliminates the need for life-span analyses until stock construction is completed. Analysis of these strains with respect to life-span are in progress. We expect that if the tester Unc locus and the Age loci are unlinked we should see 82% of such Unc he linkage would result in lower frequencies of Unc Linkage group assignments will be confirmed by the standard technique of crossing the Unc utants to wild type, reisolating F5 Unc's and testing their life-span. The age (hx546) allele prolongs life-span so dramatically that genotype determination can be reliably estimated from shoulder heights on survival curves. Therefore map distances can be estimated directly from survival curves of large populations. [See Figure 1]