Worm Breeder's Gazette 7(2): 66
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
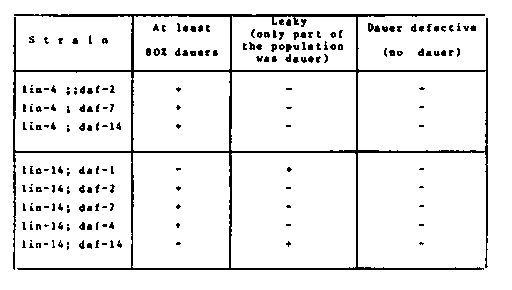
Although both lin-4(CB912) and lin-14(MT529) had originally been scored as dauer defectives, I found SDS resistant dauer larvae in starving populations of both strains. In case of lin-4 the frequency of dauers in a population seemed to depend upon the size of the original inoculum. Being interested in the interaction of lin and dauer constitutive ( daf-c) mutant genes, I constructed double mutant strains homozygous for (1) lin-14 and daf-c(daf-1(m40); 70); ) ; 72)and 7) respectively; (2) lin-4 and daf-c 370); 72) and daf-14(m77) respectively, by using szT-1 (Deak and Fodor) and C1 (R.K. Herman) balancer strains for the proper crosses. A. Construction of daf-c/daf-c; szT-1(lon-2++)/+ dpy-8 (which segregate szT-1/0;daf-c/daf-c males for crosses) was made by producing dpy-8 -c hermaphrodites and szT-1/0;daf-c/+ males from progeny of the same cross; then crossing them with each other and selecting for NonDpy NonUnc dauers at 25 C. (The procedure was a little bit different with daf-1 because of the maternal effect.). B. Construction of lin-14 szT-1/0;daf- c/daf-c males were crossed to lin-14/lin-14; m-5+ hermaphrodites. Eggs were collected at 18 C and developed at 25 C. Cross progeny could be distinguished from the self not only visually but on the basis of progeny test (Lon males segregated from cross progeny.) F1s were cloned and kept at 25 C and dauer progeny were collected, recovered, cloned and grown at 20 C. Those which did not segregate Lon males were retested for lin-14 and daf-c homozygosity by putting respectively early L4 and L1 progeny to 25 C. C. Construction of lin-4 szT-1/0; daf- c/daf-c males were crossed to C1(dpy-10+unc-52)/+unc-4+ hermaphrodites. F1 males were crossed to CB912 hermaphrodites. F1s were cloned and kept on 25 C. Those which segregated both dauers and DpyPara1 (but no Unc-4) phenotypes were kept. Dauers were recovered and cloned on 18 C. From plates which did not contain DpyPara1s, vulvaless animals were picked up, cloned and tested for the expression of the daf-c gene. All tests were repeated for several generations. 'Egg preps' were produced by the hypochlorite method (R. Hecht) from presynchronized populations. Eggs were allowed to hatch in M9 on 25 C. The L1 populations were transferred to large NGM plates spread with bacteria. Data below came from repeated experiments. [See Figure 1] The temperature-induced LinDaf dauers were SDS resistant but morphologically a little bit different from starvation dauers. It was not possible to select for higher penetrance of the daf-14 gene in the lin-14; on. In further experiments I studied the response of lin-4 and lin-14 DRIF (WBG 6, (1:) 25). Synchronous L1 populations were liquid media containing DRIF (a gift from Jim Golden) and 0.2% of bacteria and some antibiotics. Both strain seemed to grow slower than the N2 controls. After three days, practically 100% of lin-4s but less than a half of lin-14s were dauers. Some lin-14 worms seemed to be retarded. CB912 responded to DRIF normally, therefore the relatively low frequency of dauers on their starvation plates might be interpreted as a decreased production (or release) of the DRIF. MT529 responded to the DRIF in variable way. It might be interpreted that there are cells responsible for or participating in the dauer formation which cannot respond to the DRIF (or to some other DRIF-induced 'signal '), when lin-l4 is expressed in them making them biologically too old or too young to do so. However, daf-2, s seem to be epistatic over lin- 14. On the other hand, I saw lin-14; growing from L2 via dauer larvae to L4 on 25 C.