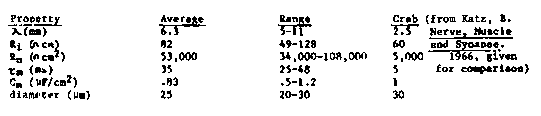
Worm Breeder's Gazette 7(2): 55
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Microscopic studies (Stretton et al., PNAS, 75:3493, 1978; White et al., Phil. Trans., 275:327, 1916) have revealed the morphological similarities between motorneuronal types found in Ascaris des and those found in C. elegans. Based on the assumption that the function of morphologically analogous elements will be similar, we have previously predicted that types DAS, DB and DA (Ascaris types DE1, DE2 and DE3) are excitatory motorneurons whereas types VD and DD (Ascaris VI and DI) are inhibitory motorneurons. At the last C. elegans meeting (1981), we described techniques which allowed us to make the first intracellular recordings from Ascaris neurons. These recordings were made from the commissures of identified motorneurons. (Commissures are single motorneuron processes which connect the ventral and dorsal nerve cords. They occur in both Ascaris and C. elegans.) At that time, we noted that spontaneously occurring excitatory and inhibitory potentials were conducted with very little decrement for long distances along the commissure (~.5 cm). Using two intracellular microelectrodes for stimulation and recording, we have now determined the electrical ( cable) properties of commissures: [See Figure 1] Commissural membranes are similar to excitable membranes found in most other organisms in two of their properties: internal (axoplasmic) resistivity (Ri) and membrane capacitance (Cm). Commissures differ rather markedly from most other excitable membranes in their space constant (lambda) and membrane resistance (Rm). We suspect that the unusually high Rm is an inherent property of the commissural membrane itself since the Cm is similar to that of a single biological membrane. (Myelinated fibers with their wrapping of multiple membranes have a much lower Cm.) We are now examining this anatomically. The high Rm produces a long lambda which accounts for the ability of the commissural membrane to conduct spontaneous passive signals over long distances with only slight decrement. We have also examined electrical signals evoked by stimulation of the ventral nerve cord. To date, all of the signals we have observed are graded, i.e., the signals lack a clear threshold and their amplitude is a continuous function of stimulus strength. We have never observed spontaneous all- or-none action potentials nor have we been able to evoke them (at normal resting potential or after hyperpolarization). Thus, lacking this classical long-distance signalling mechanism, Ascaris motorneurons appear to rely on their unusual membrane properties. to convey information over the long distances separating the two nerve cords. Nonetheless, active voltage-dependent membrane channels do appear to be present as indicated by our ability to elicit anode-break responses. These responses, however, are themselves graded (i.e., they increase in amplitude with increasing strength of the hyperpolarizing pulse). The signalling, properties of the interneurons are as yet unknown, though we are in the process of investigating them. At this point, it is not possible to rule out the presence of an action potential mechanism in other nematodes ( including C. elegans). Such an active mechanism may, however, not be necessary; in fact, one might expect passive signalling to be the modus operandi in a system where only short distance signalling (on the order of one or a few millimeters) is required. If the Rm of neuronal membranes in C. elegans is the same as that in Ascaris commissures, a lambda of 0.9mm for a 0.5 m fiber (in adult C.e.) or 0. 4mm for a 0.1 m fiber (in the L1) would be expected. Thus passive signalling over the length of the animal is entirely feasible. We have also examined neuromuscular synaptic transmission. For both excitatory and inhibitory motorneurons, synaptic transmission is graded. In addition, we have evidence that Ascaris motorneurons tonically release neurotransmitter. It is easy to see how graded synaptic transmission can be an effective way to generate the continuous gradation of muscle contraction which underlies a locomotory wave (in contrast to integration of multiple non-graded neuronal elements which together could produce the same effect). It is also tempting to speculate that graded synaptic transmission and/or tonic neurotransmitter release may be important in the production and variation of muscle tonus comprising the hydrostatic skeleton.