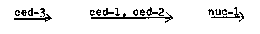
Worm Breeder's Gazette 7(2): 44
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
In the last Newsletter [7(1)] we described the identification of the gene ced-3, which appears to be necessary for the initiation of programmed cell death in C. elegans. We have constructed double mutants involving ced-3 and three previously identified genes known to affect cell deaths: ced-1 and ced-2 mutants prolong the highly refractile stage of cell death (Ed Hedgecock, personal communication), and nuc-1 mutants fail to degrade the DNA of dying cells and result in the persistence of pycnotic nuclei (John Sulston, personal communication). The epistasis of ced-1 and ced-2 over nuc-1 and of ced-3 over all three other genes suggests that these genes act sequentially in a developmental pathway for programmed cell death: [See Figure 1] We believe that this program is expressed by most cells that undergo programmed cell death, but there may be exceptions. For example, the death of the male linker cell, which requires the presence of a 'killer cell' (Sulston et al., Develop. Biol. 78: 542, 1980), can occur in ced-3 mutants. Perhaps ced-3 is required only for deaths that are cell autonomous. At least some cells that survive in ced-3 mutants differentiate. We previously noted that the postdeirids of ced-3 animals contain supernumerary dopaminergic neurons (presumably the cells that die in wild type) and that the male-specific cephalic companion neurons (the homologues of which die in wild-type hermaphrodites) appear to survive and express normal characteristics in ced-3 hermaphrodites. More recently we have obtained evidence that two additional classes of surviving cells differentiate in ced-3 animals. First, ced-3 mutants contain supernumerary pharyngeal serotonergic neurons: in ced-3 animals four pharyngeal cells have a serotonin-uptake system, whereas in wild-type animals only two neurons, known as the NSM's, accumulate exogenous serotonin (Horvitz et al., Science 216: 1012, 1982). John Sulston has found that the embryonically-generated NSM's are the sisters of cells that die in N2, which suggests that the extra serotonin cells in ced-3 animals may be the sisters of the NSM's. Second, the hermaphrodite-specific neurons (HSN's), the homologues of which die in wild-type males, appear to survive and express normal characteristics in ced-3 males. Carol Trent has recently isolated a number of Egl (egg-laying defective) mutants that lack HSN function (see Trent, Tsung and Horvitz, this Newsletter). Two of these mutants, egl-1(n487) V and n695 V, have no HSN's at hatching. Both of these mutations are dominant and are suppressed by ced-3. ced-3; lay eggs and have HSN's, suggesting that the Egl phenotype of egl-1 reflects the expression in hermaphrodites of the normally male-specific program for HSN cell death. That ced-1; odite embryos have supernumerary cell deaths when and where the HSN-homologues die in wild-type males supports this interpretation (we used ced-1 to facilitate the identification of these cells). ced-3; 5) animals have HSN's; however, other phenotypes associated with n695 (see Trent and Horvitz, this Newsletter) are not suppressed by ced-3. We are currently generating Egl+ revertants of eg1-1 to isolate additional ced-3 alleles as well as mutations in other genes that block programmed cell death.