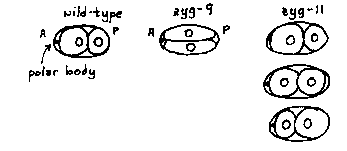
Worm Breeder's Gazette 7(2): 27
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
In the last newsletter I reported the finding of several commercial preparations of rabbit and goat IgG that fortuitously stain granules unique to the germ line (P granules) in C. elegans. Several additional antibody preparations have now been generated. From a screen of 26 rabbits from the wilds of Arvada, Colorado, the serum from 2 rabbits contains anti-P-granule antibodies. A hybridoma cell line that secretes IgG directed against P granules was obtained by immunizing mice with C. elegans egg homogenate and screening the resulting antibodies by immunofluorescence microscopy on fixed C. elegans eggs. Several new hybridomas that synthesize anti-P-granule antibodies are presently being cloned. Hopefully, at least some of these antibodies are directed against different determinants on P granules. Mutants that show defects in the early embryonic clevage patterns are being used to determine whether the position and orientation of the mitotic spindle and cleavage furrow are involved in granule segregation. A wild-type zygote divides transversely and asymmetrically. Zyg-9 (b244) zygotes divide first longitudinally and then transversely (see below). Despite the altered orientation of the spindle and cleavage furrow, P granules are localized at the posterior pole of the embryo and thus are distributed to both of the resulting blastomeres. This result suggests both that P granules do not segregate with one nucleus or one half-spindle and that the positional information responsible for P-granule segregation is separate from the positional information that orients the spindle and cleavage furrow. Zyg-11 (b2) zygotes exhibit variable positioning of the first cleavage plane, as shown below. The localization of P granules in zgy- 11 embryos is also variable and difficult to interpret. In a strain carrying mn40, an amber allele of zgy-11, a large percentage of early embryos have P granules localized at the end of the embryo next to the polar body, which in wild-type embryos is almost always anterior. It is not known whether in these zgy-11 embryos the polarity of the whole embryo is reversed relative to the polar body, or whether P granules are segregated to the wrong end of the embryo. In many of the symmetrical 2- and 4-cell zgy-11 embryos, P granules are found in all of the cells. These results support the suggestion that this mutant is defective in the establishment of polarity (see Wolf, Kemphues, Wood & Hirsh, this issue). To complement these mutational manipulations, C. elegans embryos are being treated with inhibitors of microtubule- and microfilament- mediated processes to investigate the possible roles of these cytoskeletal components in P-granule movement. [See Figure 1]