Worm Breeder's Gazette 16(3): 18 (June 1, 2000)
These abstracts should not be cited in bibliographies. Material
contained herein should be treated as personal communication and
should be cited as such only with the consent of the author.
Vulva patterning in P. pacificus:Dynamic cell migration, ring formation and cell fusion
Irina Kolotuev,
Benjamin Podbilewicz
Department of Biology, Technion-IIT, Haifa, 32000, Israel
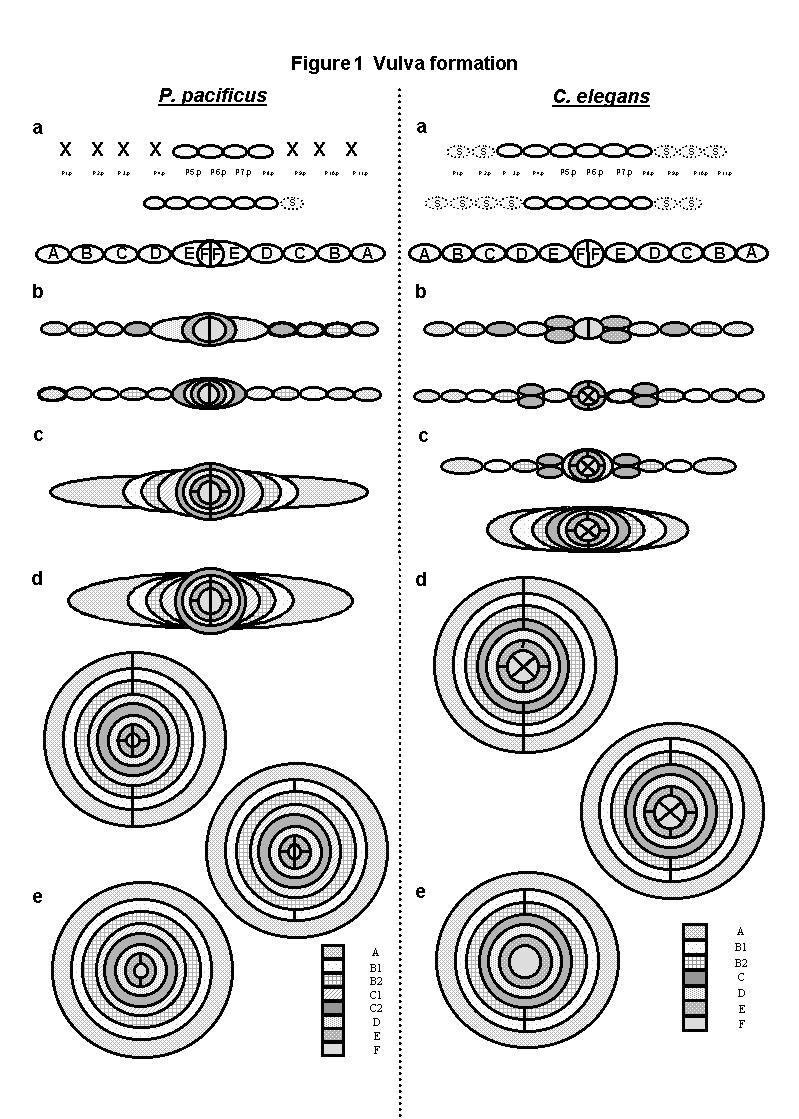
Vulva development can be observed cell by cell in different species of nematodes. In many species of the order Rhabditida it has been possible to directly compare vulval lineages. In Nematoda there are variations in the number and fates of precursor cells (Pn.p) as well as in the inductive mechanisms (1). Pristionchus pacificus (Diplogastrina) has been described as a satellite model of C. elegans (Rhabditina) for developmental biology studies. The molecular, genetic and developmental approaches to study these two nematodes separated evolutionarily by about 100 million years are very similar (2). The vulval lineages and the basic cell fate decisions have diverged in C. elegans and in P. pacificus. For example, while in C. elegans the early cell fate decision is between to fuse to the hypodermal syncytium (S) or to become vulval, in P. pacificus the decision is between whether to die (X) or to become vulval (Fig. 1a; (2)).
To study vulva formation in P. pacificus we analyzed confocal 3D-reconstructions of developing vulvae and followed the generation of vulval cells, ring formation (migration), stacking of rings and cell fusions. In P. pacificus the vulva is generated from 20 cells of the ventral epidermis in the middle of the animal, while in C. elegans the number is 22 at the same location. In C. elegans the invariant lineage of all precursor cells (LLTNTTTTNTLL. L=longitudinal, T=transverse, N=undivided) is a stage that ends before the beginning of the ring formation stage and results in the formation of a two dimensional array of cells with the fates ABCDEFFEDCBA (3). To our surprise, we found that in P. pacificus the last transverse divisions (LLLNTNNTNLLL) occur at a more advanced stage of cell migration, when there is already an invagination and after the formation of four vulva rings (Figs. 1b & 1c). During the concerted migration of vulval cells toward the central axis, eight vulva rings are formed in P. pacificus compared to seven in C. elegans (Figs. 1c-1e). The generation of an additional ring in P. pacificus is the result of a L division in C, that in C. elegans normally fuse transversely at the beginning of morphogenesis, but they do not fuse in P. pacificus and give rise to two separate rings. One of these rings (C2) is the first one to undergo intratoroidal fusion even before the end of other cell migrations (Fig. 1d). In the last stage of vulva formation, the intratoroidal fusions, in P. pacificus follow the sequence C2, C1, D, A/B2, B1, F and E compared to D, A, C, F, and E in C. elegans (Figs. 1d & 1e).
By comparing the morphogenesis of rings in vulvae from two species it can be seen that homologous rings do not only differ with respect to the exact structures that they produce, but also behave quite differently with respect to the time-course of their formation and fusion (heterochrony). The mechanistic and evolutionary significance of this data will probably require a full comparison of morphogenetic pathways between different nematode species.
1. M. A. Felix, et al., Dev Biol 221, 68-86 (2000). 2. A. Eizinger, B. Jungblut, R. F. Sommer, Trends Genet. 15, 197-202 (1999). 3. R. Sharma-Kishore, J. G. White, E. Southgate, B. Podbilewicz, Development 126, 691-699 (1999).