Worm Breeder's Gazette 13(4): 54 (October 1, 1994)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
| 1 | Biology, Indiana University, Bloomington, IN 47405. |
| 2 | current address: Molecular Biology, University of Oregon, Eugene, OR. |
| 3 | Zoologisches Institut der Universitat Koln, Kerpener Str. 15, 50923 Koln, Germany |
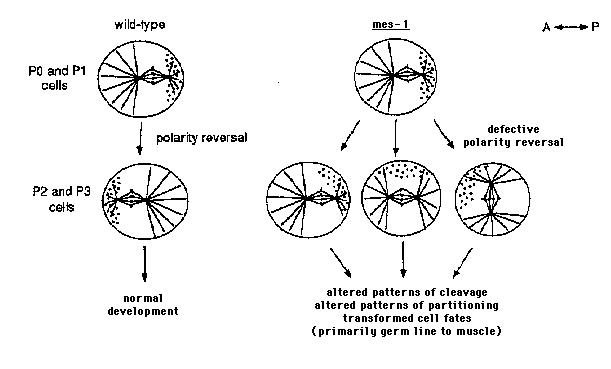
The mes-1 gene was identified in screens for maternal-effect sterile mutations, which cause homozygous mutant mothers to produce sterile progeny (Capowski et al. Genetics 129, 1061-1072, 1991). mes-1 mutant progeny are sterile because they fail to form primordial germ cells during embryogenesis. The primary defect in mes-1 embryos appears to be in the process of polarity reversal in P2 .Defects in polarity reversal lead to altered patterns of cleavage and cytoplasmic partitioning and to transformations in cell fate, primarily a transformation of the germ-line founder cell, P4 ,into a muscle precursor, like its sister, D. Our results are summarized below. (see figure) All 10 mes-1 alleles are both temperature sensitive and incompletely expressed. Homozygous mutant mothers generally produce 10-20% sterile progeny at the permissive temperature (16260C) and 60-80% sterile progeny at the restrictive temperature (25260C) (Capowski et al., 1991). This is probably the null phenotype, since mes-1 mutations behave similarly to a deficiency of the region. Thus, even in the absence of wild-type mes-1 product, the process in which the gene participates can be executed normally, especially at low temperature. The earliest and probably primary defect in mes-1 embryos is in polarity reversal in P2 .In wild-type embryos, a series of unequal P-cell divisions set the germ line apart from the somatic lineages. In wild-type embryos released from the constraints of the eggshell (i.e. "partial embryos"), P1 and P2 are generated to the posterior and P3 and P4 to the anterior of their somatic sisters. None of the 23 partial mes-1 embryos examined at 25260C displayed normal polarity reversal. Instead, the divisions of P2 and P3 displayed a variety of deviations from the normal pattern; for example, some divisions were oriented along the A-P axis but were relatively symmetric and others were asymmetric but oriented transverse to the A-P axis. These division patterns reflect mis-positioning of the spindle in the P2 and P3 cells of mes-1 embryos. mes-1 embryos display defects in the divisions of P2 , P3 ,and P4 .In intact mes-1 embryos, the timing of divisions and sizes of blastomeres are normal until the 7-cell stage, after which they differ from wild-type embryos in the following ways: 1) The divisions of P2 and P3 do not display the pronounced asymmetry seen in wild-type embryos. 2) P4 often divides precociously and undergoes extra rounds of division, at about the same time as its somatic sister, D. Embryos that display precocious and supernumerary divisions of P4 hatch into L1 sthat lack the germ-line cells Z2 and Z3 and develop into sterile adults. Thus, the sterility of mes-1 animals is due to defects in the divisions that normally generate the primordial germ cell, P4 .Fertile mes-1 adults arise from embryos that undergo a normal lineage. mes-1 embryos display defects in cytoplasmic partitioning. P granules are correctly partitioned to P1 and to P2 in mes-1 embryos, but are mis-partitioned during the divisions of P2 and P3 .For example, in the 16- to 46-cell embryos examined, 18% contained P granules in only P4 (as seen in wild type), 80% contained P granules in both P4 and D, and 2% contained P granules in D alone. The cause of the mis-partitioning appears to be that P granules are often segregated to the side of the spindle in P2 and P3 ,instead of to the pole of the spindle destined for the P-cell daughter (as in wild type). On consequence of mis-partitioning is that P granules are often located in ectopic cells (probably muscle) in mes-1 larvae. In mes-1 embryos P4 is transformed into a muscle precursor. Our lineage analysis suggested that in at least some mes-1 embryos P4 follows the fate of its sister, D, and generates body muscle instead of germ line. Two results support this germ line to muscle transformation hypothesis. First, sterile mes-1 adults contain up to 20 extra body muscle cells, the number normally generated by D. Using a transgenic muscle reporter construct from Andy Fire, sterile mes-1 worms contain 103 261 7.8 muscle nuclei compared to 94.7 261 1.5 in wild-type worms and 94.6 261 1.9 in fertile mes-1 worms. Second, "isolated" P4 blastomeres in mes-1 embryos generate muscle cells. In laser-operated wildtype embryos (see Bowerman et al. Cell 74, 443-452, 1992 for procedure), isolated D blastomeres generate muscle and isolated P4 blastomeres do not, as judged by the ability of the cells to contract and by staining with anti-myosin antibody (Ab from David Miller). In contrast, in mes-1 embryos almost all (18/19) isolated P4 blastomeres produced muscle cells; D blastomeres also produced muscle. Thus, in mes-1 embryos, P4 cells contain the potential to generate muscle and are most likely responsible for generating the extra body muscle cells observed in sterile mes- 1 adults. Summary and model. We hypothesize that either the program or the machinery underlying polarity reversal is inherently sensitive to temperature, and that wild-type mes-1 product guarantees the successful execution of polarity reversal at both high and low temperature. This insures that the progeny of P2 (C, D, and P4 )have the correct sizes, contents, and neighbors, and that they follow their normal fates. In the absence of mes-1 product, there is loss of coordinate regulation of spindle positioning and cytoplasmic partitioning in P2 and P3 (see below), leading to progeny cells with altered sizes, contents, and developmental programs. The main developmental defect is a transformation of the germ-line cell, P4 ,into a muscle precursor, like its sister. D.