Worm Breeder's Gazette 13(4): 50 (October 1, 1994)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Department of MCD Biology, University of Colorado, Boulder
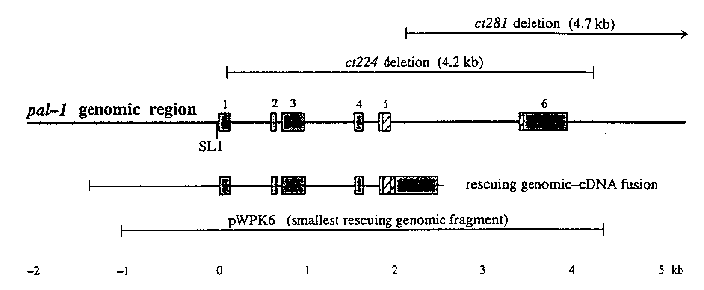
We have previously reported that two small deletion mutations, ct224 and ct281 ,result in an embryonic lethal Nob phenotype, characterized by defective L1 'sthat hatch with a severely deformed posterior(1). These mutations fail to complement two previously described nonlethal pal-1 mutations that result in a postembryonic ray transformation in the male tail (Mab phenotype)(2). Using one of the Mab mutants, Waring and Kenyon cloned the pal-1 gene, sequenced a cDNA, and showed that the predicted protein included a homeodomain similar to that of the Drosophila caudal gene product(3). From sequencing of the pal-1 region in N2 (recently confirmed and extended by the Sequencing Project) and genomic fragments obtained by PCR from strains carrying ct224 and ct281 ,we have determined the gene structure and the extents of the two deletions (see Figure). Both deletion mutants can be rescued by the genomic-cDNA fusion construct shown, indicating that the Nob phenotype results from loss of pal-1 information rather than from deletion of an adjacent gene sequence. On Northern blots, a pal-1 probe recognizes two major transcripts of 1.25 and 1.45 kb, which are present at high levels in embryos and gravid adults, and at low levels during larval development. We are currently exploring the following questions: 1) Is maternal as well as embryonic expression of pal-1 required for embryonic development? When marked pal-1 / qC1 hermaphrodites are injected with the rescuing cDNA or genomic constructs shown in the figure, the rescued pal-1 homozygous progeny are infertile, producing only Nob embryos in the next generation. Fertile pal-1 homozygous lines have so far been obtained only by injection and integration of the entire pal-1 cosmid. One explanation for these results could be that there is a maternal requirement for pal-1 expression, and that the injected DNA is expressed poorly in the germ line. Consistent with this view is the finding that pal-1 mRNAs are present at high levels in adult fem-1 and fer-1 (no fertilized embryos) but absent in glp-4 ( bn2 )hermaphrodites, showing that they are restricted to the germ line in adult hermaphrodites. 2) Is Nob the null phenotype and Mab a hypomorphic phenotype for pal-1 ?Over the small deficiency rhDf1 ,kindly provided by Kiyoji Nishiwaki, which deletes pal-1 and the flanking genes emb-5 and dpy-17 ,(4)both ct224 and ct281 result in Nob phenotypes similar to those of the corresponding pal-1 homozygotes, suggesting that this is the null phenotype. Strains homozygous for the viable Mab allele pal-1 ( e2091 )produce about 5% Nob embryos, and 30% to 50% of the L1 'sshow a bulge near the middle of the animal which disappears in later larval stages. However, e2091 / rhDf1 animals produce no higher frequency of Nob embryos and show the L1 bulge and Mab phenotypes characteristic of e2091 homozygotes. This result suggests that the Mab mutants are not simply hypomorphic pal-1 alleles, but rather that they may define a separate pal-1 function in male tail development which is not required in the embryo. 3) How does the Pal-1 protein function in posterior patterning? We previously reported expression of a pal-1 -LacZ reporter construct in a small number of cells which arise in or migrate to the posterior of the embryo, (l) and we have since extended these experiments to include several similar constructs containing different amounts of pal-1 upstream and coding sequence. We find that all constructs containing at least 0.5 kb of upstream sequence and coding sequence corresponding to the first three exons result in production of up to 10% Nob embryos in an N2 background, whereas constructs with less coding sequence do not. These results suggest that an overproduced amino-terminal PAL-1 fragment interferes with normal posterior patterning, perhaps through deletenous interactions with other proteins that normally complex with Pal-1 .This possibility would be consistent with recent evidence that Drosophila ftz and other homeodomain proteins participate in protein-protein interactions that do not require the homeodomain(5). It would also predict possible differences in phenotypic effects of the two deletions ( ct224 in fact results in a slightly more severe Nob phenotype than ct281 ),which we are currently analyzing in more detail. Understanding pal-1 embryonic function will clearly require more detailed knowledge of the Nob phenotype. With the help of Eric Putz in this department, we have (finally) succeeded in building a "4D" digital video recording system based on Andy Fire's design (personal communication) and adapting his software to a Leitz DMR Nomarski microscope, which has the advantage of an electronically controlled focusing system that can be directly driven by the computer (details on request). Analysis of the embryonic lineage in Nob embryos from video recordings is just beginning. (see figure) (1) Edgar et al. (1993) C. elegans Meeting Abstracts, p 10. (2) Waring and Kenyon (1990) Cell 60: 123-131. (3) Waring and Kenyon (1991) Nature 350:712-715. (4) Described in Ahnn and Fire (1994) Genetics 137:483-498. (5) Brown and Hartley (1994) Nature 370:414 415.