Worm Breeder's Gazette 13(4): 33 (October 1, 1994)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
| 1 | Department of Biology, New York University |
| 2 | Department of Molecular Genetics, Albert Einstein College of Medicine |
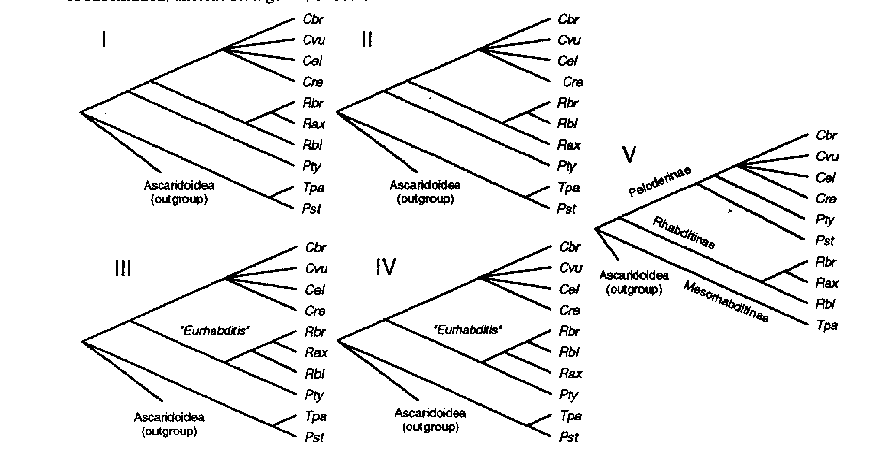
We have investigated the molecular evolution of the nucleotide sequences of 18S ribosomal RNA genes (18S rDNA) from part of our collection of nematodes in the family Rhabditidae. Our aim was to evaluate the usefulness of this gene for molecular systematics of this family, as well as to establish phylogenetic relationships within a group that has potential for comparative studies of the relationship between development and evolution. We determined the 18S rDNA sequences of 9 species of nematodes representing 6 genera within this family: Caenorhabditis briggsae (Cbr), Caenorhabditis vulgaris (Cvu), Caenorhabditis remanei (Cre), Rhabditis blumi (Rbl), Rhabditis sp. br (Rbr), Rhabdilella axei (Rax), Pellioditis typica (Pty), Teratorhabditis palmarum (Tpa) and Pelodera strongyloides dermatitica (Pst). Using hypothetical models for secondary structure as well as nucleotide similarity, these sequences were aligned with the 18S rDNA sequence published for C. elegans (Ellis et al. 1986, Nucl. Acids Res. 14:2345-2364) and with the partial sequences published for 8 ascaridoid species (Nadler 1992, Mol. Biol. Evol. 9:932-944). We found that 18S rDNA is likely to be a useful tool to resolve relationships at the intrafamilial level. However, 18S rDNA sequences cannot be used (by themselves) to resolve relationships between taxa as closely related as the Caenorhabditis species. Parsimony, minimum evolution and maximum likelihood methods strongly reject one proposed phylogenetic classification based on adult morphological characters (Andrassy 1983, tree V below), but support that of Sudhaus (1976, tree IV) as one alternative of a few possible phylogenies (trees I-IV) that were significantly better than all other topologies tested. When variation in substitution rate over different sites is modeled by a gamma distribution, there is stronger support for monophyly of Rhabditis than when rate differences are ignored. When transversions are weighted as phylogenetically more informative than transitions, or only conserved positions are considered (2/3 of the total data), support jumps markedly for monophyly of the "Eurhabditis" clade ( Rhabditis species, Rhabditella and Pellioditis ). Based on these trends in the data, we predict that 18S data from additional taxa will support a tree (i.e., tree IV below) very similar to that proposed by Sudhaus (1976). Distances between 18S rDNA sequences of different Caenorhabditis species (0.8-1.8%) are comparable to distances between 18S rDNA of different orders in class Mammalia. Distances between genera in Rhabditidae are about 8 times as great as distances between tetrapod classes, suggesting rapid rates of substitution, ancient divergence, or both. (see figure) Andrassy, 1983. A taxonomic review of the suborder Rhabditina (Nematoda:Secernentia). ORSTOM, Paris Sudhaus, 1976. Vergleichende Untersuchungen zur Phylogenie, Systematik... Zoologica 43(125): 1-229