Worm Breeder's Gazette 13(3): 92 (June 1, 1994)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
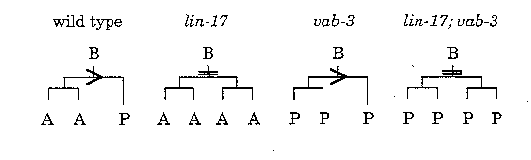
The initial division of the C. elegans male B cell is asymmetric: the anterior daughter of B (B.a) is the precursor to all of the cells of the copulatory spicules, whereas B.p produces no spicule fates (Sulston, et al. (1980) Dev. Biol. 78, 542-576). In addition, B.a is larger, divides earlier, and produces more progeny than B.p. lin-17 is required for the initial unequal division of the B cell as well as for fate specification (Sternberg and Horvitz, (1988) Dev. Biol. 130, 67-73). In lin-17 mutants the initial division of B is equal, and both B.a and B.p behave like a normal B.a cell. In genetic screens for mutants with male B lineage defects we recovered two vab-3 mutations ( sy66 and sy281 ;e.g., WBG 11(3), pg. 56). vab-3 has many functions in development (A. Chisholm, WBG 11(4), pg. 83). We have found that in the B lineage vab-3 uncouples the unequal cell division from fate specification. In vab-3 mutants, B divides unequally, and B.a divides to produce a left and a right daughter as in wild type. However, lineages in two vab-3 ( sy66 )and six vab-3 ( e648 )males suggest that both B.al and B.ar behave like a normal B.p cell (although there is some lineage variability in the mutants). In addition to a transformation of the axis and timing of divisions of the B.a(l/r) cells some differentiated fates are also transformed. B.ppaa normally undergoes a programmed cell death, and in the vab-3 mutants the corresponding cells in the B.a(l/r) lineages can also die. Thus vab-3 is necessary for normal B.a or B.a(l/r) fates.
We constructed lin-17 ; vab-3 double mutants. While the lineage defect in vab-3 mutants results in a distinctive pattern of six progeny in the L3 larval stage rather than the normal ten progeny, in lin-17 mutants there can be sixteen. In lin-17 ( n677 ); vab-3 ( e648 )males there can be eight progeny. The defects are additive: the initial division of B is equal because lin-17 is mutant, but both B.a and B.p behave like the abnormal B.a of vab-3 mutants. Thus lin-17 might be necessary for both the unequal cell division and the asymmetric distribution of fate potential, whereas vab-3 acts downstream to specify B.a or B.a(l/r) fate. In contrast, Mike Herman's analysis of lin-44 and lin-17 suggests that lin-44 acts upstream of lin-17 in the B lineage (Herman and Horvitz (1994) Dev. in press).
Lineage analysis of lin-17 ; vab-3 double mutants suggests that lin-17 may have additional functions in the B lineage. Specifically, predicted B.p like lineages are abnormal, and the cells generally divide synchronously (two animals followed). To identify additional functions of lin-17 in the B.a and B.p lineages, we followed the B lineage in four lin-17 mutants that had escaped the initial defect at the division of B, and the B.a lineage in two lin-17 animals in which B.p was ablated. The results suggest 1) lin-17 functions in the B.p lineage to mediate the asymmetric division of B.pp, 2) lin-17 functions in the B.a lineage to mediate the asymmetric cell division of B.a(l/r)ap and possibly B.a(l/r)pa, and 3) lin-17 does not appear to play a role in the divisions of B.a(l/r) or B.a(l/r)(a/p). Thus lin-17 functions at multiple, but not all, asymmetric cell divisions in the B lineage.
Interpretation of B cell lineage defects in lin-17 , vab-3 ,and lin-17 ; vab-3 mutants