Worm Breeder's Gazette 13(3): 74 (June 1, 1994)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
In the C. elegans hermaphrodite the seal between the intestinal lumen and the exterior is maintained by structural constraints of the anal region. Contraction of specialized enteric muscles, most importantly the anal depressor, is required to break this seal and permit defecation. The neurotransmitter GABA appears to be the major excitatory transmitter that activates this contraction. A large number of mutations have been isolated that cause a defect in anal depressor contraction, and hence a Constipated (Con) phenotype (Thomas, 1990). However, analysis of males mutant for most these genes revealed that they are not Con, suggesting that male defecation occurs by a mechanism different from that used in the hermaphrodite.
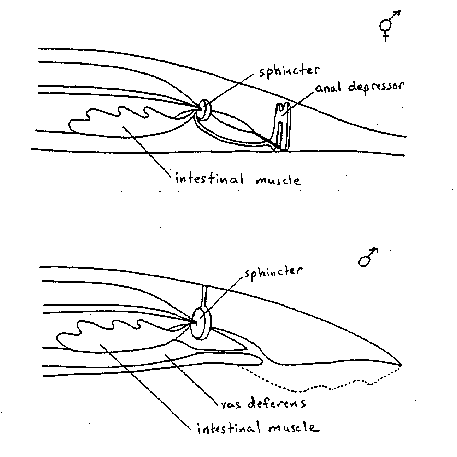
A comparison of the structure of the hermaphrodite anal region and that of the male revealed several interesting differences (1, 2). While the hermaphrodite anus is intrinsically sealed, the male anus is an open cloaca. The spicules, used during mating, are contained in the cloaca In addition to the intestine emptying into the cloaca, the vas deferens transfers sperm through this canal during mating. Near the end of the L4 the male anal muscles undergo a dramatic reorganization. The anal depressor reorients to become a spicule retractor, and the previously minor (and functionally insignificant) anal sphincter hypertrophies and forms a dorsal attachment. The sphincter has been proposed to function in the male for sperm release by contracting to open the vas deferens ( 1). Here we propose that the sphincter also forms the seal between the intestine and the cloaca, perhaps by tonic partial contraction. This would predict that relaxation of the sphincter would be necessary for defecation, and that hyper-contraction results in sperm release.
Analysis of mutant males defective in anal depressor contraction revealed that most of them did not have a Con phenotype. The exceptions are mutants in unc-25 and unc-47 ,genes thought to be involved in GABA synthesis and vesicle loading, respectivtely (3). These males are severely Con. Since we hypothesize that no muscle contraction is required for defecation in the male, why are unc-25 and unc-47 males Con? The answer is revealed by observation of mutants in unc-46 and unc-49 ,which are implicated in inhibitory GABA neurotransmission. These mutant animals are also defective in male defecation, albeit somewhat less severely than unc-25 or unc-47 males. These observations suggest a model in which GABA is used as an inhibitory neurotransmitter in the male to relax the sphincter during defecation. Males defective in inhibitory GABA neurotransmission are defective in sphincter relaxation, and are therefore constipated.
We tested this model by killing the male sphincter muscle with the laser in unc-25 , unc-47 and unc-46 mutant backgrounds ( unc-49 was not included in this analysis because of its weaker male Con phenotype). Killing the sphincter rescues the male Con phenotype for each gene. In addition, the kill caused constitutive leaking from the anus in mutant and wild-type males. We also showed that the inhibitory GABA agonist muscimol was also capable of rescuing the mutant male Con phenotype. These results indicate that tonic sphincter contraction seals the male anus, and that GABA is required to relax the sphincter.
We also wished to know which neurons are responsible for male defecation. The GABAergic motomeurons AVL and DVB are required for anal depressor contraction in the hermaphrodite (3), and we have demonstrated that they are similarly required in the larval male. However, AVL, DVB operated males are not Con as adults. Since unc-25 males are Con as adults, we propose that there are other GABAergic motomeuron(s) required for relaxation of the sphincter in the adult male. Killing of obvious candidate cells has proved fruitless, and further experiments' await identification of GABAergic neurons in the male.
Finally, we addressed the question of unc-25 function, since this gene is central to our analysis. Recent work has shown that unc-25 encodes the GABA biosynthetic enzyme GAD (4). We used a temperature-sensitive (ts) unc-25 mutation isolated by Dennis Liu to address whether GABA is required during development for later function. If unc-25 were required in a developmental process, it would presumably have a distinct ts period. We showed that unc-25 ts mutants do not have a distinct ts period. Instead, function can be partially removed or restored by t-shift regardless of developmental stage. In addition to the rescue of unc-25 excitatory function by exogenously added GABA (3), we demonstrated that exogenous GABA can rescue the male sphincter-relaxation defect of unc-25 .These results are consistent with unc-25 encoding a biosynthetic enzyme.
This system is ripe for genetic analysis. Screens for Con adult males could identify genes necessary for the development or function of the GABAergic neurons used to relax the adult sphincter. Screens for suppressors of the unc-25 adult male Con phenotype could yield genes necessary for sphincter development or function, or for neurons necessary for tonic sphincter contraction. This system provides additional opportunity for study of general GABA function in the worm nervous system as well as the development and nature of a sexually dimorphic behavior.
(2) White, et al. (1986) Phil Trans R Soc Lond B 314: 1-340.
(3) McIntire, et al. (1993) Nature 364: 334-337 and 337-341.
(4) Jin, et al. (1994) WBG 13(2): 69.