Worm Breeder's Gazette 13(3): 60 (June 1, 1994)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
The homeotic selector (HOM-C) genes are used to specify region-specific fates in many organisms. In C. elegans the HOM-C genes lin-39 , mab-5 ,and egl-5 are expressed specifically in the regions in which they are required;(1) loss of or ectopic expression leads to transformation of region-specific fates.(2) The polyray-1 ( pry-1 )gene is required to keep C. elegans HOM-C genes repressed outside of their normal domains of expression (WBG 12(5):39). In pry-1 ( mu38 )animals, the initial expression of the HOM-C genes is correct, but as development proceeds these genes are ectopically expressed, leading to dramatic homeotic transformations. mu38 is recessive, so it likely represents a loss-of-function allele. The wild-type function of pry-1 is, therefore, to maintain HOM-C genes in a repressed state once a pattern of expression has been initiated. The Polycomb-Group of genes in Drosophila serve an almost identical function to that of polyray: once the cascade of early regulators has established an expression pattern of HOM-C genes, the Polycomb genes are required to maintain genes in a repressed state where they weren't originally expressed.(3)
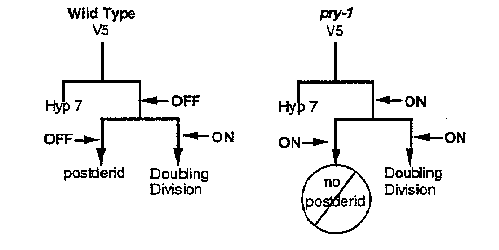
The classification of genes which regulate HOM-C expression as either initiators that establish pattern or maintenance genes that stably maintain an established pattern is appealing but perhaps simplistic. The pattern of HOM-C expression is dynamic, even after the "maintenance" phase begins, both in Drosophila(4) and C. elegans. How are the changes in expression accomplished? Either the maintenance system does not act in those cells which change their expression patterns, or the maintenance system does act but is modified to allow for changes in HOM-C expression. In the V5 lineage the second method appears to be used. In a wild-type worm, mab-5 is not expressed in the V5 lineage until V5 .ppis born (see Salser and Kenyon, this WBG). In fact, mab-5 must be kept OFF in V5 .paso that a postderid can be mate, but it must be expressed in the V5 .pplineage to produce a L3 doubling division (Salser and Kenyon, this WBG). In pry-1 mutants, mab-5 expression begins in V5 .p;as a result 75% of pry-1 animals develop without a postderid. The loss of the postderid is due to ectopic mab-5 ,because in pry-1 ; mab-5 double mutants, all animals make a postderid. Thus in wild-type, pry-1 is needed to keep mab-5 repressed in V5 .pand V5 .pa; pry-1 activity must then be overcome to allow for expression of mab-5 in V5 .pp.This demonstrates that temporal patterns of expression can be created by modifying the activity of a gene whose major function is maintaining patterned expression. We are planning to look for mutations that prevent mab-5 from being expressed in V5 .pp,with the hopes that these will lead to a better understanding of how the maintenance function of pry-1 is regulated.
(2) See Salser and Kenyon (1994), TlG 10:159-164.
(3) Reviewed by Paro (1990). TlG 6: 416-421.
(4) Karch et al (1990). Genes and Development 4:1573-1587. Carroll et al.(1986). Cell 47:113-122.