Worm Breeder's Gazette 13(3): 44 (June 1, 1994)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Germ cells in C. elegans must decide among a number of possible cell fates: they can act as a stem cell and undergo another mitotic cell division or they enter meiosis and undergo gametogenesis, in which case they have to decide between spermatogenesis and oogenesis. How these decisions are regulated is the subject of intense investigation (e.g. see review by Schedl, Curr. Opin. Gen. Dev, 1, 185, 1991). We have undertaken the study of another germ cell fate: programmed cell death. Our results suggest that programmed cell death is a quantitatively important cell fate in the germ line and occurs extensively as a part of or as a by-product of oogenesis.
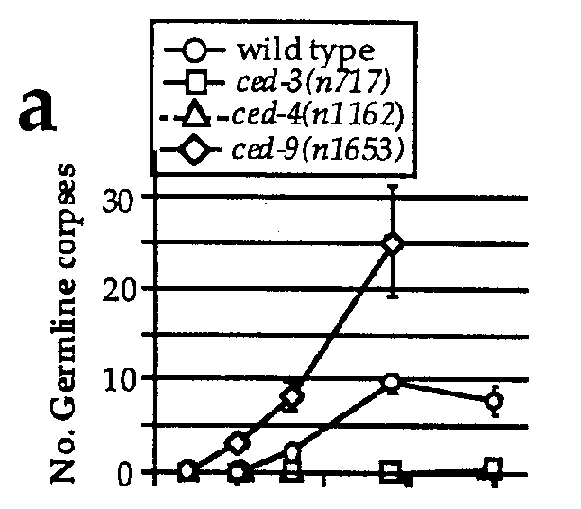
To determine the extent of programmed cell death in the germ line, we determined the number of germ cell corpses visible at various stages of development. We found that adult hermaphrodites, but not larvae or adult males, had in their germ line a significant number of refractile bodies highly reminiscent of somatic cells undergoing programmed cell death (Figure 1a), suggesting that these structures are dying germ cells.
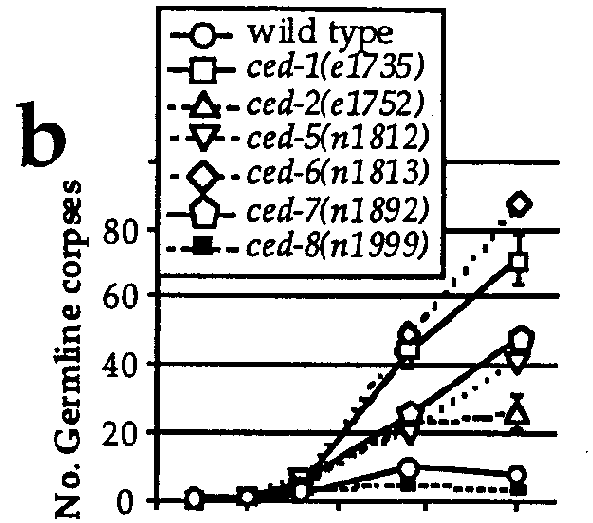
A number of genes have been described that function in the somatic programmed cell deaths (for reviews see Ellis et al., Ann. Rev. Cell Biol. 7, 663, 1991 and Driscoll, J. Neurobiol. 23, 1327,1992). We found that most, but not all, mutations that affect somatic cell deaths also affect germ cell deaths. For example, the genes ced-1 ,2,5,6,7,and 10, which function in the engulfment of dying somatic cells, are also required for efficient engulfment of dying germ cells by the sheath cell (Figure 1b). We confirmed that the refractile bodies that were accumulating in the germ line of these mutants were indeed unengulfed cell corpses by analyzing thin sections of mutant gonads using electron microscopy (EM). Many typically "apoptotic" nuclei could be seen. All dead cells observed had separated from the common germ line syncytium, suggesting that cellularization is part of the germ cell death program. Dying cells could also be observed in EM sections of wild-type gonads, but at a much lower frequency. Interestingly, we found that ced-8 mutants had fewer germ cell corpses than wild-type animals (Figure 1c), suggesting that ced-8 is not required for efficient engulfment but rather might be involved in some other aspect of germ cell death. Indeed, these results and a reevaluation of the ced-8 phenotype in embryos have led to the suggestion that ced-8 is required either for the execution or timing of programmed cell deaths, both in the germ line and the soma (see abstract by Stanfield et al. in this issue).
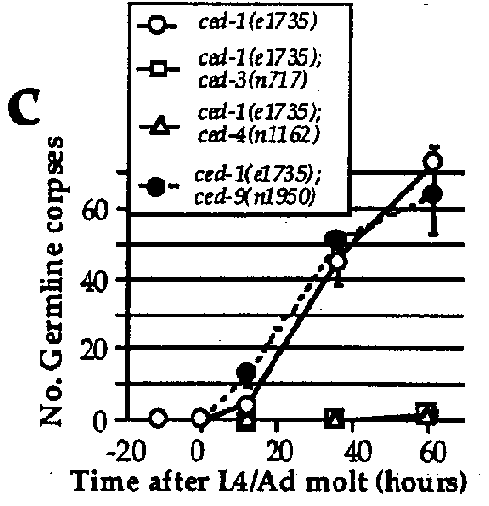
The genes ced-3 and ced-4 ,required for somatic programmed cell deaths, are also required for the programmed death of germ cells: essentially no corpses could be observed in the germ line of ced-3 or ced-4 mutants (Figure 1a). Similarly, ced-9 ,which is required to protect somatic cells that should live from programmed cell death, also protects germ cells from programmed death: in ced-9 (If)animals, many more germ cells undergo programmed cell death than in the wild type (Figure 1a), possibly explaining the partial sterility associated with the loss of ced-9 function. Surprisingly, the gain-of-function mutation ced-9 ( n1950 ),which completely blocks somatic programmed cell death, has little or no effect on germ cell death: ced-1 ; ced-9 double mutants have approximately the same number of germ cell corpses as ced-1 single mutants (Figure 1c). The n1950 mutation results in a G-to-E substitution in a domain highly conserved with other members of the ced-9 / bcl-2 family (Hengartner and Horvitz, Nature, in press). Yin et al. (Nature, in press) have shown that this conserved domain is important in Bcl-2 for heterodimerization with Bax, another member of this gene family (Oltvai et al., Cell 74, 609, 1993). The involvement of this region in Bcl-2 /Baxheterodimerization suggests that n1950 might affect interaction of CED-9 with other, unidentified proteins. One model consistent with our results is that CED-9 interacts with two partners, one in the soma and the other in the germ line. n1950 might affect interaction with the somatic partner but not the germ line-specific partner. Alternatively, unlike the situation in the soma, ced-9 might be necessary but not sufficient to prevent programmed germ cell deaths. Analysis of germ cell death in strains that overexpress wild-type CED-9 protein should help resolve this issue.
Several lines of evidence suggest that the extensive programmed cell deaths we observe are a part or a by-product of oogenesis. (1)The first germ cell deaths appear shortly after the switch form spermatogenesis to oogenesis. Even in ced-9 (lf)mutants where most presumptive oocytes die, mitotic proliferation (and spermatogenesis) appear superficially normal. (2)No cell deaths are observed in the mitotic part of the germline. The deaths are clustered around the bend in the gonad, where oocytes form. (3)Males, which make no oocytes, have little if any germ cell corpses under normal conditions. (4)In gld-1 mutants, germ cells cannot undergo gametogenesis. We observed little if any programmed cell death in gld-1 ( q268 )hermaphrodites.
What is the function of these deaths? Our current hypothesis is that cell death is used to remove excess presumptive oocytes. The germ cells, in addition to forming gametes, also function as nurse cells to direct the synthesis of the high levels of cytoplasmic components that each oocyte incorporates when it cellularizes. It is possible that the number of nurse cells required exceeds the number of oocytes that can be made, suggesting that the excess germ cells that cannot become oocytes are discarded by causing them to die. From the number of germ cell corpses that accumulate in the engulfment mutants, we estimate that at least one third (and probably significantly more) of all presumptive oocytes undergo programmed cell death.
We will screen for mutants that affect this process in the hope of understanding how the decision between oogenesis and death is regulated. We are also investigating how environmental signals affect germ cell deaths: John Sulston's observation of a high incidence of cell death in the germline of aged or starved animals (Worm Book, p.147) suggests that additional pathways can lead to programmed cell death in this tissue.