Worm Breeder's Gazette 13(1): 72 (October 1, 1993)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Three tissues have been implicated in vulval development: the anchor cell, the hyp7 syncytial hypodermis, and the Pn.p cells. We have used the ncl-1 clone in mosaic analysis experiments to determine the site(s) of action for two genes involved in specifying vulval cell fates, lin-31 and mpk-1 .
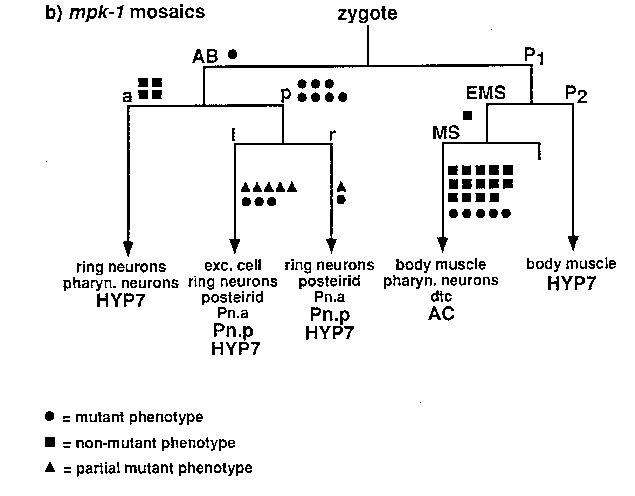
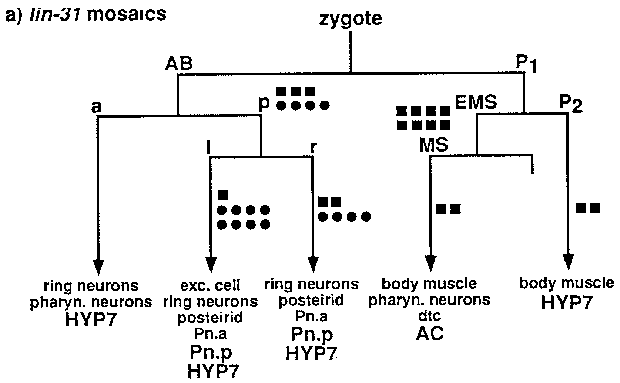
For our mosaic analysis experiments, we have used mitotic loss of an extrachromosomal array carrying two mosaic marker genes ( ncl-1 and unc-29 )and our gene of interest. Use of an array circumvents the need for free duplications and the requirement that genes of interest and mosaic markers be linked and was made possible by the identification of the cosmid ( C33C3 )that contains ncl-1 (D. Waring, WBG 12:5:16). ncl-1 mutations, which cause many cells in the worm to have enlarged nucleoli, allow single-cell resolution in mosaic analysis experiments by scoring the Ncl phenotypes of diagnostic cells to determine the point in the cell lineage at which the array was lost. unc-29 ,which encodes a subunit of the acetylcholine receptor, is also a good mosaic marker. As expected, its site of action appears to be in body muscles. Screening for Semi-Unc animals provides an easy way to identify losses on the P1 side of the lineage, since all but one body muscle derive from that side of the lineage. Semi-Unc animals can be readily identified as animals that are Unc on a plate but thrash in liquid.
Several observations argue against the potential problem that the observation of a Ncl phenotype might reflect poor expression from the array rather than loss of the array. First, cells closely related in the cell lineage, such as the Pn.a descendants or the cells that make up the postdeirid, almost always display the same Ncl phenotype. Second, most mosaic animals can be explained by losses of the array at one, or perhaps two, places in the lineage; we did not observe mosaic animals in which the pattern of Ncl cells was derived from random points in the cell lineage. Third, the Ncl and Unc-29 phenotypes correlate with losses in the same cells.
We used our new mosaic technique to determine if lin-31 acts in the Pn.p cells. Iin-31 is an HNF-3 /forkhead transcription factor homolog. In lin-31 mutants, each Pn.p cell can adopt any one of the three possible cell fates (Miller et al, 1993, Genes and Dev., 7:933). Animals of genotype unc-29 ( e1072 ); lin-31 ( n1 O53); ncl-1 ( e1875 )were injected with C45D10 ( unc-29 (+)), pLM268 ( lin-31 (+)),and C33C3 ( ncl-1 (+))to generate a non-Unc, non-Muv, non-Ncl transgenic strain. 2,600 animals from this strain were screened for Semi-Unc mosaics or using Nomarski optics for Ncl mosaics. Preliminary results are shown in Figure (a). Each square or circle represents one mosaic animal. The position of each symbol indicates the point in the lineage at which the array was lost. The results implicate the Pn.p cells as the site of action for lin-31 ,since loss of lin-31 (+)DNA in the Pn.p cells results in a lin-31 mutant phenotype (variable Muv), while loss in the anchor cell does not. Because hyp7 derives from several places in the lineage, these data are not consistent with a site of action in hyp7 .If that were the case, one might expect partial mutant phenotypes in mosaic animals. As expected, we observed that not all Pn.p(-) mosaics displayed a Muv phenotype (84%, n= l9 ),since the lin-31 ( n1 O53)phenotype itself is incompletely penetrant (80% of n1 O53animals are Muv).
A similar strategy for mosaic analysis was used for mpk-1 ,a C. elegans MAP kinase homolog and a suppressor of the let-60 (gf)Muv phenotype (see Lackner et al, this issue). Thus far, 4,500 animals have been screened to generate the data in Figure (b). The Pn.p cells are also implicated as the site of action for mpk-1 .Since the array showed incomplete rescue of the mpk-1 phenotype (23% of animals carrying an array still suppress the let-60 gfphenotype), it was not surprising that 26% (n= l9 )of the Pn.p(+) AC(-) mosaic animals still displayed a mutant phenotype. Thus, our data suggests that both a putative signal transducer ( mpk-1 )and a putative transcription factor ( lin-31 )act in the receiving cells during vulval induction, indicating that these genes are involved in the cellular response to an inductive signal.