Worm Breeder's Gazette 12(5): 62 (February 1, 1993)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Over the past year or so we have been trying to find a tool that would allow us to study the electrical properties of pharyngeal cells. We believe that we have found a technique that can be used to study some of these properties.
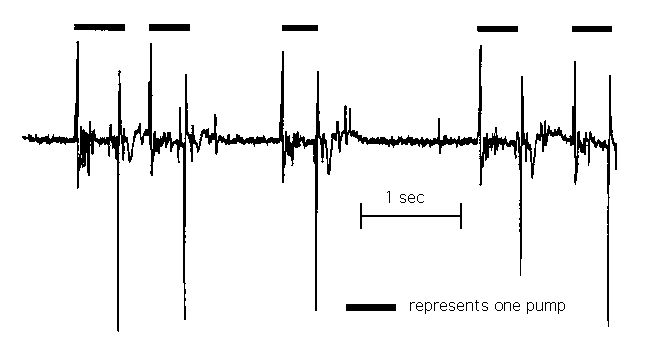
We use a differential amplification extracellular recording arrangement with the signal electrode in the pipette and the indifferent electrode wrapped around the pipette. We suck a worm's head into the pipettes and begin recording. Figure 1 shows an example of a recording from a pumping worm. We call the signal that corresponds to one pump (see figure 2) an Electropharyngeogram or EPG. The invariant features of the EPG are an initial positive spike followed by a larger, negative spike about 200 msec later. 10-30 msec before the positive spike, there is usually a smaller positive spike and 20-100 msec after the large negative spike, there is often a smaller, negative spike and then a slower negative deflection of about 100 msec duration. Between the two large spikes there are often several small, negative deflections. (The overshoots after the large positive and negative spikes are an artifact of high pass filtering.) There is a perfect correlation between the electrical signals and pharyngeal motions. The beginning of corpus contraction is perceived within 67 msec after the positive spike, and corpus relaxation occurs within 33 msec after the negative spike. We believe that at least the two main spikes are caused by electrical changes in the pharynx and not by motion artifacts because a) the positive spike occurs before the first perceptible motion in the pharynx and the negative spike occurs at the same time or before relaxation is perceived and b) EPGs recorded from act-2 ( ad468 ),an actin mutant with greatly attenuated pharyngeal motions, are of the same shape and roughly the same amplitude as the signals from wild type.
What is the origin of the EPG? The two large spikes are likely of muscle and not neural origin for three reasons. First, the correlation between pharygeal muscle motions and those spikes is perfect; we never see one without the other. Second, there are thought to be few spiking neurons in nematodes [Davis and Stretton J Neurosci 9, p. 403]. Third, with an AC-coupled amplifier, faster changes in potentials result in larger signals recorded. Relaxation of the pharynx, which is faster than contraction [Avery, J Exp Biol 93, in press], is correlated with a larger spike than the one correlated with contraction. A rigorous test for the origin of the EPGs will be to record from a worm after ablating its pharyngeal nervous system. The origin of the small, negative deflections between the two large spikes is a mystery. Interestingly, some of the action potentials recorded from Ascaris pharyngeal muscles cells also have negative deflections during the plateau phase of the action potential [Del Castillo and Morales, J Gen Phys 50,1967, p. 603]. Generally similar records to ours were obtained from the parasitic nematode Ancylostoma caninum [Roche et al, Science 136, 1961, p 148] but their 3 published records did not appear to have these deflections.
The polarity of the EPG is informative. The signal electrode is in electrical continuity with the pharyngeal lumen. If the luminal surface of pharyngeal muscle cells were electrically active during the action potential, the spikes associated with the start and end of the action potential would have opposite polarities from those observed; hence, we suspect that the active ion channels are found only on the basolateral surface of pharyngeal muscle cells and that the signal is coupled to the lumen passively through the luminal membrane. This makes biological sense since the ionic composition of the worm's environment is variable and generally not conducive to the production of action potentials.
Because the shape and duration of the EPG are fairly reproducible between worms and between pumps, we can easily detect gross abnormalities in the signal. Indeed, preliminary experiments suggest that three mutants with abnormal pharyngeal motions, eat 4, eat-G, and eat-12 ,have EPGs that are different from wild type. We hope to determine whether there are electrical abnormalities in Eat mutants with no obvious pharyngeal motion defects. Since the time between the positive and negative spike corresponds to a single pharyngeal pump one can use the electrical recording to measure pump length over long periods of time. We recorded over 1000 EPGs from a single N2 worm in the presence of serotonin and found the pump length to be 184.6 + 1.1 msec (mean SEM). Such high precision may allow us to discern small changes in pump length in mutants, laser-ablated worms, and in response to drugs.
[See Figures 1&2;]
