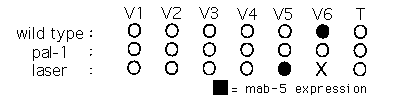
Worm Breeder's Gazette 12(5): 37 (February 1, 1993)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Generation of a normal pattern of sensory rays in the C. elegans male tail involves activity of the genes mab-5 and pal-1 ,as well as signaling between the lateral epidermal cells. Based on genetic studies, pal-1 and cell-signals appear to act as upstream regulators of mab-5 ,which, in turn, promotes ray production. Using polydonal antibodies against mab-5 we have confirmed this model and shown that regulation of mab-5 occurs through control of its expression.
The Pal-1 ( e2091 )phenotype(transformation of V6 into a V4 -1ikecell producing alae instead of rays) results from apparent reduction-of-function mutations in a gene required for embryonic development. Although the nature of these reduction-of-function mutations is still unclear, pal-1 (+)has been shown to be required autonomously, in V6 ,for production of rays. The requirement for pal-1 can be overcome if mab-5 is provided using the gain-of-function allele e1751 ,suggesting that pal-1 may simply be needed for mab-5 expression in V6 .Using polyclonal antibodies to visualize mab-5 we have found that pal-1 is indeed required for mab-5 expression in V6 .Whereas wild-type L1 larvae almost always express detectable levels of mab-5 in V6 (76/78), pal-1 ( e2091 )mutant larvae almost never do (1/40). Expression in QL, M, body muscle, and juvenile motor neurons is apparently unaffected. V5 descendants do not normally express mab-5 until L2 .This expression is presumeably unaffacted in pal-1 ( e2091 ),for these animals produce a normal V5 lineage complete with its one ray.
Ablation of V6 allows V5 to produce up to flve rays as opposed to its normal complement of a single ray, a postdeirid, and an adult seam celL Production of these rays by V5 is known to require mab-5 ,suggesting that activation of ray production may proceed through activation of mab-5 .Using antibodies we have found that mab-5 expression indeed switches on in V5 .pfollowing ablation of V6 . V6 was ablated on one side of each animal within 30 minutes of hatching. 12 hours later, animals were fixed and stained for mab-5 expression. V5 .pon unablated control sides never expressed detectable levels of mab-5 (0/14), whereas V5 .pon the ablated side sometimes expressed mab-5 (3/14). Staining efficiency in this experiment was rather low, (V6.p expressed in only 5/13 sides) perhaps due to difficulty in handling small numbers of larvae, or due to variations in expression levels during late L1 .However, we believe this result is significant, as in other control experiments, expression has never been seen in V5 descendants until L2 .
These experiments demonstrate that cell-signalling interactions repress mab-5 expression, and that pal-1 is needed to permit mab-5 expression in V6 .They do not distinguish whether pal-1 acts directly to promote mab-5 transcription, or whether it acts to antagonize repressive cell signals. Assuming the second model is correct, then repressive cell signals must be active prior to hatching, for newly hatched pal-1 ) animals do not express mab-5 in V6 .
[See Figure]