Worm Breeder's Gazette 12(5): 23 (February 1, 1993)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Transposition of the Tc3 transposon has been detected in the high
hopper strain TR679 which was initially selected for increased
transposition of the related element Tc1 .Both Tc1 and Tc3 insert at
a TA dinucleotide. A consensus sequence around this TA dinucleotide
has been proposed for Tc1 insertions in the germ line (G A G/T A/G
T A C/T G/T T; Mori et al., 1988, Proc. Natl. Acad. Sci. USA 85:86t-
864; Eide and Anderson, 1988, Mol. Cel. Biol. 8:737-746); this
consensus sequences is based on the comparison of 16 Tc1
insertions. We have sequenced 96 independent somatic Tc3
insertions to determine the Tc3 integration consensus sequence.
This is part of an ongoing study in which we compare Tc1 and Tc3
integration site specificity and in which we investigate the
predictive value of the Tc1 integration consensus sequence
mentioned above.
In the previous WBG (WBG, 1992 12/4:15) we described a transgenic
strain in which we could induce the expression of Tc3 transposase in
somatic tissues. The transgenic strain was made in a Bristol N2
background in which no Tc3 transposition occurs. After induction,
frequent somatic Tc3 transposition events were observed, as
witnessed by the appearance of extrachromosomal Tc3 elements and
by insertions of Tc3 in a random gene (the gpa-2 gene). Insertions
were detected by two rounds of PCR with nested oligo's. To
determine the insertion sites, the PCR products were excised from
an agarose gel and sequenced according to Craxton (Methods: A
companion to methods in enzymology, 1991 3:20-26).
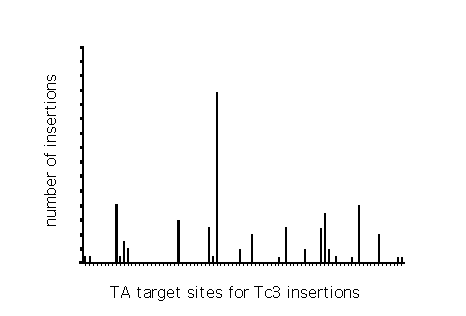
The distribution of Tc3 insertions in an 1 kbp region of gpa-2 is
depicted in the figure. Each small line on the X axis represents a TA
dinucleotide: a potential Tc3 insertion site. The vertical bars
represent the number of Tc3 insertions found at a specific TA
dinucleotide.
[See Figure]
From the distribution we conclude that Tc3 insertions are not
random. TA dinucleotides which are frequently used for integration
are not clustered or regularly spaced. Furthermore, the TA
dinucleotide with the most frequent Tc3 insertion (24/96) is
flanked by two TA dinucleotides which are used only 1/96 or 0/96
times. This suggests that the local rather than the regional
structure of the DNA facilitates insertions.
The alignment (according to the 5'-3' orientation of the gpa-2 gene)
of all the 96 insertion sites of Tc3 results in the consensus
sequence: A(40%)/T(44%) A(60%) A(49%)/C(41%) T A T(79%) T(70%)
A(44%)/T(53%). The consensus sequence is not very strict; it does
not resemble the Tc1 consensus sequence.