Worm Breeder's Gazette 12(4): 60 (October 1, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
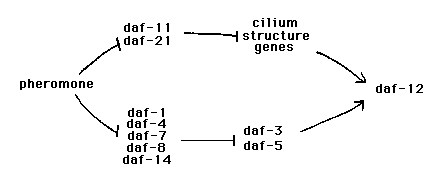
The decision to enter the dauer larva stage is controlled by sensory cues, primarily the concentration of the dauer pheromone in the environment. Most of the genes implicated in control of dauer formation fall into two classes, those with dauer formation constitutive mutations (Daf-c) and those with dauer-formation defective mutations (Daf-d). These genes have been ordered by conventional epistasis analysis into a formal genetic pathway. We are interested in dauer formation as a problem in processing of chemosensory information, and thus our attention is largely focused on the early steps in this complex pathway, which we designate arbitrarily as those upstream of daf-12 .The players in the comedy, then, are: the Daf-c genes daf-1 ,4, 7, 8,11,14, 21, and the Daf-d genes daf-3 , daf-5 , daf-6 ,and a host of genes required for normal amphid sensory cilia, which we call collectively the cilium-structure genes.
Until recently, nearly all gene interactions among these upstream genes have appeared consistent with a simple linear pathway. Nevertheless, certain results left us with nagging doubts about this simplicity. First, all the Daf-c genes in this part of the pathway are represented only by incompletely penetrant alleles. These alleles are Ts, probably because of the intrinsic temperature-sensitivity of dauer formation (see Malone and Thomas in this WBG, and Golden and Riddle, PNAS 81: 819). Second, each of these Daf-c mutants responds strongly to dauer pheromone at low temperatures, indicating that some functional transduction pathway remains intact. Third, the one daf-3 mutation and two daf-5 mutations previously tested strongly suppress the Daf-c phenotype of daf-1 ,4, 7, 8, and 14, but poorly suppress daf-11 and 21. This third result conflicts with the fact that mutations in the cilium-structure genes strongly suppress daf-11 and 21 mutations but show no suppression of daf-1 ,4, 7, 8, and 14.
All of these results can be explained away, without invoking a nonlinear pathway, by the usual suspects (non-null alleles, allele-specific daf-3 and daf-5 interactions etc.). However, recently we have obtained new information that further damages the linear model. We constructed the full matrix of daf-c; daf-c double mutants among these upstream genes. A striking pattern emerged. Recall that all of these Daf-c mutations are incompletely penetrant. In most double mutants the Daf-c phenotype was similar to that of the stronger single mutant. However, certain pairs of double mutants displayed a powerful synergy: all double mutants between daf-11 or 21 (henceforth "group 1" genes) and daf-1 ,4, 7, 8, or 14 ("group 2" genes) form 100% dauers at any temperature. This result suggests that there is partial redundancy between two processes involving these two groups of genes. Remarkably, these two groups of genes correspond exactly to the groups as defined previously by epistasis analysis: the cilium-structure Daf-d mutations are epistatic to mutations in the group 1 genes, but not the group 2 genes
These results suggest that there are two parallel processes interpreting sensory cues. Normally both processes are integrated to determine whether to form a dauer, but activation of either process alone can partially induce dauer formation. We have also made other observations that support this interpretation. First, we have extended the analysis of daf-3 and daf-5 by isolating three new daf-5 mutations and seven new daf-3 mutations, and by more extensively testing their epistasis with respect to the group 1 and group 2 Daf-c genes. This analysis shows that the strong suppression of group 2 but not group 1 genes is characteristic of all of these daf-3 and daf-5 alleles. We hypothesize that daf-3 and daf-5 act only on the group 2 branch of the pathway, downstream of the group 2 Daf-c genes. Second, we have tested pheromone responsiveness of both daf-c single mutants and daf-c; daf-c double mutants within each group at 15°. This analysis is not quite complete, but all single and double mutants so far tested retain a clear response to pheromone. We hypothesize that the Daf-c mutations activate one of the parallel pathways but leave the other pathway functional, thus accounting for their pheromone responsiveness.
Two other features of the dauer pathway fit neatly with this model. First, we have found that daf-12 mutations are able to fully block the activation of both parallel sensory processes, suggesting that daf-12 acts at or downstream of the convergence of the parallel sensory pathways. Second, daf-2 is the only gene for which fully penetrant Daf-c mutations exist (D. Riddle, pers. comm.). daf-2 is epistatic to both the cilium-structure mutations and mutations in daf-3 and daf-5 ,and has a lethal interaction with daf-12 (Vowels and Thomas, Genetics 130: 105). These facts suggest that daf-2 also acts at or downstream of the convergence of the parallel sensory pathways. [See Figure]
Vowels and Thomas, Genetics 130: 105.