Worm Breeder's Gazette 12(4): 54 (October 1, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Pharyngeal muscle is capable of rhythmical contractions (pumps) without a nervous system (Avery and Horvitz Neuron 3:473). The function of pharyngeal neurons is to modulate the rate and rhythm of pharyngeal pumping in response to the environment. How does the environment control pharyngeal pumping rate?
We know at least one environmental condition that affects pumping: food. When worms are transferred from an environment without food to one with food, their rate of pharyngeal pumping increases. We also know at least one of the neurotransmitters that regulate pumping rate: serotonin. External application of serotonin stimulates pumping. Finally, we know that there is a serotonergic neuron(s) active in the absence of food. Imipramine, a serotonin-uptake inhibitor, increases the rate of pumping in the absence of food in wild-type but not in the serotonin-deficient mutant, cat-4 (Avery and Horvitz J Exp Zoo 253:263). What is the endogenous source of serotonin that is enhanced by imipramine?
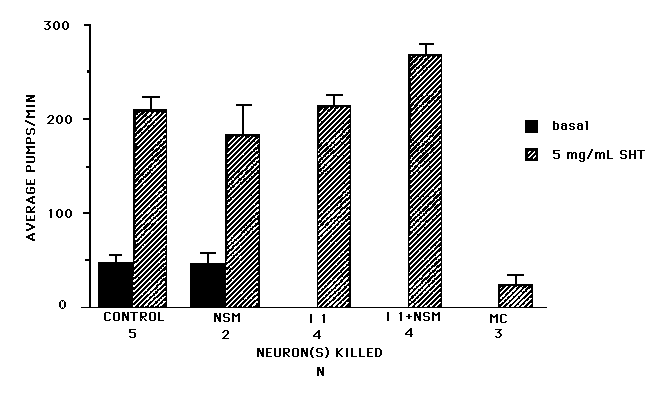
There are two plausible candidate sources for this serotonin: RIH and the NSMs. NSM is the only pharyngeal neuron type known to contain serotonin, and RIH is an extrapharyngeal serotonergic neuron with indirect inputs onto the pharyngeal neuron types I1 and M1 .To see if one or more of these neurons is necessary for the basal rate of pumping and for the stimulatory effect of imipramine, we killed the three pharyngeal neuron types I1 , M1 ,and NSM. Indeed, in NSM-, I1 -, M1 -worms, basal pumping is almost completely abolished and there is no stimulatory effect to imipramine (20 mg/ml, [See Figure 2]). We then killed subsets of these three neurons. Neither NSM nor M1 kills alone had a significant effect on the basal rate of pumping or on imipramine stimulation. Killing I1 alone or together with NSM resulted in an obvious phenotype. I1 -worms seldom pumped in the absence of food with or without imipramine [See Figure 2].
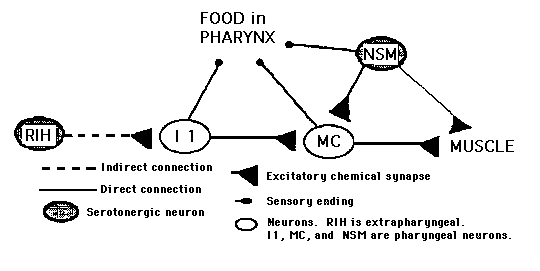
Figure 1 represents our current view of the rate-control circuit. I1 has output onto MC, a sensorimotor neuron type that is known to be necessary for the increased pumping rate in response to food. Hence, it is likely that I1 is excitatory onto MC which, in turn, is excitatory onto pharyngeal muscle. I1 is the major recipient of input from the extrapharyngeal nervous system and has an indirect connection with RIH, a serotonergic neuron. Therefore, we suggest that in the absence of food, RIH activity dictates the rate of pumping via I1 .Imipramine enhances the effect of serotonin secreted by RIH only in the presence of I1 .
Surprisingly, exogenous serotonin stimulates pumping in worms that lack I1 [See Figure 3] Is serotonin bypassing I1 and acting directly onto MC to stimulate pumping? Consistent with this possibility, we find that the response of MC- worms to exogenous serotonin is attenuated [See Figure 3]. An alternative possibility is that serotonin does not affect MC directly but that MC is partially necessary for the response to serotonin. At the moment, we cannot distinguish between these possibilities. Serotonin may have a small MC-independent effect on pumping rate, in addition to the larger MC-dependent effect [See Figure 3].
We propose that there are two neural circuits that control the pumping rate, a pharyngeal one and an extrapharyngeal one [See Figure 1]. The pharyngeal circuit exerts the main control on rate. In response to food, pharyngeal neurons can increase the pumping rate up to ten-fold. However, they can only do so if they taste the food in the pharyngeal lumen. In order to get food into the lumen, a basal rate of pumping is necessary. This rate is controlled by input from the extrapharyngeal nervous system. According to this scheme, the pair of NSM neurons secrete serotonin only when activated by bacteria in the lumen. Therefore, NSM- worms are not different from intact worms in basal pumping or in the response to imipramine. In the presence of food, NSM could be redundant with other pharyngeal neurons. In addition to NSM, I1 and MC and six other neuron types have endings that could be mechanosensory (Albertson and Thomson Phil Trans R Soc 257: 299). Therefore, NSM- worms respond normally to food (Avery and Horvitz Neuron 3: 473).
Avery and Horvitz J Exp Zoo 253:263.
Albertson and Thomson, Phil Trans R Soc 257: 299.