Worm Breeder's Gazette 12(4): 48 (October 1, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
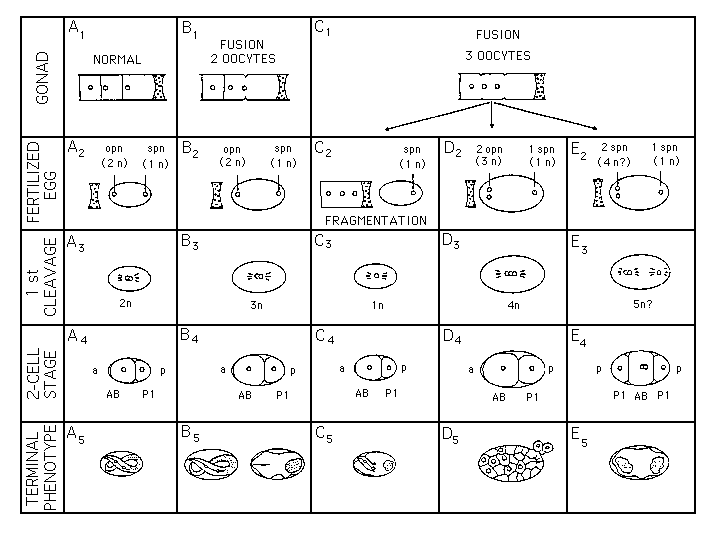
One question we are interested in, is, whether a fixed polarity is already present in the unfertilized egg of the nematode as described for other systems. In order to investigate this question we have developed a technique to fuse oocytes with a laser microbeam. Generally, the most distal oocytes (positioned closest to the spermatheka) are suited best for such a manipulation. Giant oocytes generated this way can be fertilized. So far, we have succeeded to obtain giant embryos derived from a fusion of 2 or of 3 oocytes. Not all fused oocytes become fertilized or initiate cleavage. So far, in 37 cases giant zygotes derived from 2 oocytes were found to start cleavage. All of them expressed a normal early cleavage pattern with the generation of 6 lineages. The vast majority of such embryos arrested as monsters, most of them with signs of tissue-specific differentiation However, 20% (8/37) developed into normal-looking giant juveniles [See Figure 1, B1 -B5].Most of these (6/8) reached the adult stage with essentially normal properties but having approximately twice the volume of a regular worm They reproduced with a low (15% of normal) brood size (reduced number of eggs produced; frequent embryonic arrest) with the fraction of males much higher than normal (up to 30%). During subsequent generations the number of males returned to the normal percentage (ca 0.2%). These results show that the uncleaved egg can regulate to form a (nearly) normal animal from 2 oocytes They indicate that no fixed polarity and prelocalization of cytoplasmic determinants are present in the oocyte. The high number of males and low fecundity appears to be due to triploidy and aneuploidy as observed by others (see Bob Herman, worm book).
After fusion of 3 oocytes (C1) we observed 3 different pathways of development (C-E). In two thirds of the cases (23/33) the giant oocyte was fragmented into a smaller anucleate and a larger triploid part. The anucleate fragment could be fertilized and develop as an apparently haploid embryo with a normal early pattern to eventually arrest during late embryogenesis (C2-C5), often quite normal morphogenesis in the head region (10/23) and terminal cell differentiation (muscle twitching, gut birefringence). The triploid part usually lysed during the passage through the spermatheka In no case it started cleavage.
Alternatively, such a super-giant oocyte could be fertilized in total and initiate normal early cleavage with the complete series of unequal germline divisions (5/33). So far, in all observed cases of this type embryos arrested before the beginning of gastrulation (< 30 cells) and often lysed, apparently due to a weak eggshell ( D2 -D5).Despite the arrest, these findings show that even a zygote derived from 3 oocytes can regulate to behave like a normal egg.
Some eggs derived from 3 oocytes behaved quite differently (E2-E5). Pronuclei (a single one at one pole, two at the other pole) approached each other from the poles and both formed a cleavage spindle along the anterior-posterior axis of the egg resulting in 2 small cells at the ends and a large cell in the center (5/33). With the next division the large central cell formed a tetrapolar spindle and generated 4 blastomeres of similar size and future AB-behavior. The two small cells at the poles performed a series of germline-like, unequal cleavages like a P, cell Our interpretation that this represents a development of twins fused at their anterior (AB) ends is supported by the generation of 2 distinct E-cell groups with gut-specific birefringence (ES).
So far, we consider 2 explanations for this unexpected finding: a the maternal centrosome has not been inactivated in these eggs and forms a second cleavage spindle together with the centrosome introduced by the sperm. However, so far no case has been reported that cleavage can be initiated in unfertilized eggs of C.e. or after removal of the sperm centrosome (Schierenberg and Wood; Dev. Biol 107, 337, 1985). Additionally, it is difficult to understand why a posterior pole is established under these conditions where normally the anterior pole would be. Therefore, we favor explanation b.: Because of the large egg size the assumed polyspermy-block initiated by the entrance of a sperm at the pole entering the spermatheka first is not completed (fast enough?) allowing the entrance of a second sperm at the other pole. If this assumption is true, it could mean that it is the entrance of a sperm which generates a posterior pole.
Presently, we try to test whether a second sperm has been introduced in this type of egg If anyone has a smart idea (e.g. in-vivo marking of sperm) please contact us (E-mai} address: ESchier@Biolan Uni-Koeln De; FAX: 049-221-470-3109). Experiments to fuse oocytes of giant nematodes to generate even larger worms and fuse their oocytes etc. are under consideration. Donations of giant petri dishes are welcome.
Schierenberg and Wood; Dev. Biol 107, 337, 1985.