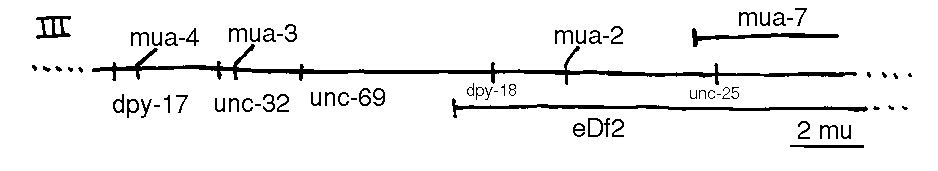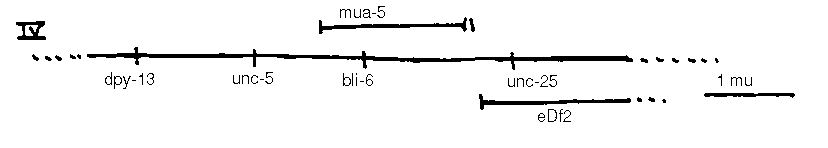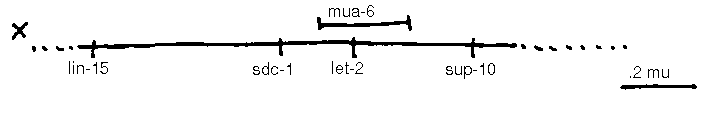
Worm Breeder's Gazette 12(3): 98 (June 15, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Normal locomotion of C elegans results from the transmission of contractile tension produced by the body wall muscles through the plasma membrane, extracellular matrix, and hypodermis to the cuticle via specialized attachment sites. In order to identify genes likely to encode molecules involved in the adherence and transmission of tension between the muscles and the cuticle we have collected mutations that cause a progressive flaccid paralysis of all or part of the body due to detachment of body wall muscles from the cuticle during larval development. These mua (for muscle attachment) mutations fall into at least 12 complementation groups and appear to comprise two broad classes. The first class appears to primarily affect the integrity of the hypodermis; the second class appears to primarily affect attachments between muscle cells and/or muscle cells and the hypodermis.
mua-3 III is an example of the first class. Observations of larvae in the process of detachment suggest that the defect may be due to an inability of the mutants to reform the hypodermal/cuticle links during the molting process. The hypodermis being too weak to withstand the tension placed on it by the muscles herniates; the muscle bands, which remain intact, collapse away from the cuticle into the pseudocoelom, sweeping hypodermal debris with them.
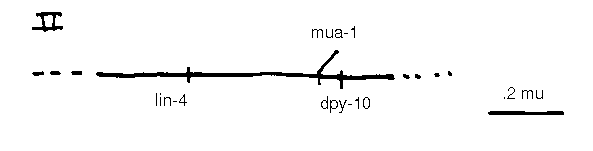
mua-1 II is an example of the second class of mua. Animals homozygous for the sing allele rh160 show progressive development of breaks between muscle cells in the muscle bands, and detachment of the bands from the hypodermis. The progression of paralysis is relatively slow, 63% of L2 ,39% of L4 ,and 5% of young adults are grossly unaffected. Electron micrographs of relatively non-paralyzed homozygous L4 larvae reveal dense bodies, normally found only at muscle cell/hypodermal attachment points, to be ectopically positioned between adjacent muscle cells. Assuming this mispositioning to be a cause rather than consequence of mua-1 muscle detachment, we suggest that the ectopic dense bodies occur at the expense of normal muscle/muscle attachments (dense plaques) weakening the muscle bands, and allowing eventual band breakage At a grosser level, rh1601 Df animals do not appear more severely affected, suggesting that rh160 may represent the null phenotype of mua-1 .We are screening for additional alleles. The intriguing phenotype, coupled with its well defined map position, have prompted us to begin the cloning of mua-1 .
In addition to the loci shown, the following loci are known: vab-10 I, mua( rh197 )II, mua( rh214 )III, unc-23 V, him-4 X. mua-5 complements bli-6 ,m ua 6 complements let-2 [See Figures 1-4].