Worm Breeder's Gazette 12(3): 43 (June 15, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
We are investigating embryonic functions of the pal-1 homeobox gene (Waring and Kenyon, Cell 69:123; Nature 350:712). The two psoralen-induced alleles ct224 and ct281 produce hatching L1 swith a a severely deformed posterior, which we call a Nob (no back end) phenotype. The earliest detectable abnormality is a misarrangement of the gut cells at the 100-cell (16 E cell) stage. Neither allele complements pal-1 ( e2091 )and we presume that they represent null alleles of this gene.
1. Using Southern analysis, we have determined that both of these alleles are deletions covering the 3' end of pal-1 . ct224 ,Approximately 4.2 kb, lies entirely within the smallest pal-1 rescuing plasmid, the 6 kb Kpn I/ Pst-1 restriction fragment pWPK6 (a gift from Waring and Kenyon), beginning in the homeobox and deleting the rest of the 3' coding sequence. ct281 is approximately 4.7 kb, beginning slightly 3' of ct224 and deleting into the next 3' Kpn 1 fragment; it may include a small part of the homeobox. Northerns show a doublet of 1.25 and 1.45 kb (the appropriate size for pal-1 )when probed with any of the pal-1 coding fragments from pWPK6 .The 3' 3 kb Kpn I fragment encodes a transcript of 800 bp which is presumably affected in ct281 but should not be affected in ct224 unless its untranscribed sequence extends beyond the Kpn I site.
2. We have injected pWPK6 ,the larger plasmids pWK9 and pWK14 ,and the cosmid W O5E6 (all gifts from Waring and Kenyon), together with rol-6 DNA, into ct224 and ct281 ,and now have integrated or transmitting lines from each possible combination. In all cases, we recover Unc Dpy animals as well as the heterozygotes from the balanced strains mut Unc Dpy/ qC1 carrying an array. However, the Unc Dpy animals produce only nonhatching embryos with a severe Nob phenotype. Although one explanation for this maternal effect is that a second, maternal effect mutation is closely linked to the deletion mutations, both alleles and the heterozygotes ( ct224 unc-32 dpy-18 / ct281 dpy-18 unc-25 )show this effect when rescued, even with the cosmid. Thus we suspect that pal-1 may also have a maternal function which is not rescued possibly because this transforming DNA is not expressed in the germ line.
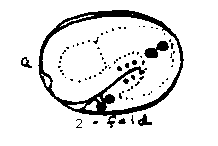
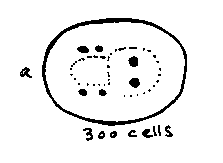
3. To investigate expression patterns of the wild type pal-1 gene, we have injected a construct containing approximately 5 kb of pal-1 upstream genomic sequence and about half the coding sequence fused to the reporter bacterial lacZ gene. The earliest expression observed is in the 4, then 8, C cells in 70-80 cell embryos. This disappears, and at approximately 300 cells Earpp, Ealpp, and 4 cells lateral to the pharyngeal primordium stain. These cells migrate posteriorly, and are probably Z1 , Z4 ,M, and mu int. By lima bean stage, several additional ventral midline cells and 2 cells at the forming rectum, probably U and K, begin to stain, continuing through hatching [See Figure 1 &2;]. Earpp and Ealpp form int-5 at the center of the gut, where Ea and Ep descendants interdigitate, and like Z1 and Z4 ,contact Z2 and Z3 during embryogenesis. U and K form the intestino-rectal junction. Thus all the staining cells lie posterior, at positions where malfunction might well give the Nob phenotype of the mutants.
Isolated and cultured P1 ,EMS, and P2 blastomeres appear cell-autonomous for pal-1 expression; isolated AB blastomeres give no blue cells. Since U and K are AB descendants, inductive interactions may be required for their expression of pal-1 .
Waring and Kenyon, Nature 350:712