Worm Breeder's Gazette 12(3): 29 (June 15, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Genetic analysis has demonstrated that tra-1 activity is necessary and sufficient for female somatic development, and that, conversely, its inactivity is required for male development. tra-1 is the terminal gene in the sex determination cascade, lying immediately downstream of the fem genes. The fem genes are thought to inhibit tra-1 activity in X0 animals, thus permitting male development. In their absence tra-1 directs feminization of XO animals (and XX germline). The phenotype of tra-1 gain of function mutations is very similar to that of fem loss of function mutations. This suggests that these tra-1 gf mutations render tra-1 insensitive to fem regulation. Analysis of the mutations causing these gf phenotypes might thus be expected to elucidate how fem regulation of tra-1 is achieved. Molecular analysis reveals that tra-1 produces two transcripts as a result of alternative RNA processing (D.Z. and J.H.). The smaller transcript encodes a protein containing two zinc finger motifs, the larger transcript a protein having five zinc finger motifs. The predicted two zinc finger protein is almost completely contained in the five zinc finger protein. The mRNA levels for each transcript are very similar in XX and XO animals. The three fem genes have also been cloned, and sequence is available for fem-1 and fem-3 . fem-1 contains six copies of the ANK repeat, a motif thought to mediate protein/protein interactions. fem-3 is a novel putatively intercellular protein (Ahringer et al, EMBO, in press).
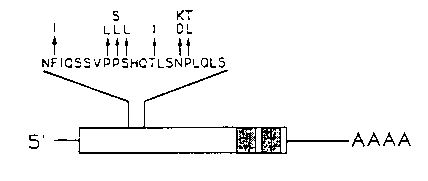
We have sequenced 14 tra-1 gain of function mutations of various strengths, ranging from Class 1 alleles, which almost completely feminize XO animals, to Class 5 which only feminize XO germline and gonad. The mutations in all these alleles cause non-conservative amino-acid substitution within a sixteen amino-acid residue region at the amino-terminal end of both the predicted tra-1 proteins as diagrammed below [See Figures]. This suggests that negative regulation of tra-1 activity occurs by a protein/protein interaction, possibly between pTra- 1 and one or more of the pFems, and that the tra-1 gf mutations identify a site required for this interaction to occur [See Figure 1].
These results engender several interesting questions. Which, if any, of the pFems binds to pTra-1? Does the gf defined N-terminal region suffice for binding the negatively regulating activity? And if this is the case, can one of the functions of the predicted small protein be to protect the larger protein from the negatively regulating activity? Most important, how might this predicted binding activity prevent pTra- 1 action in X0 animals? Does it affect nuclear/cytoplasmic localization of pTra-1, does it alter or abolish its DNA binding, or does the interaction change the activity of pTra-1 as a transcriptional activator/repressor?