Worm Breeder's Gazette 12(3): 24 (June 15, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
Comparing physical and genetic maps
Chakravarti (Genomics, 1991) has suggested that a powerful way to compare the genetic and physical maps of chromosomes would be using so called Marey maps, after the French graphic artist of a century ago. In this method, each gene has Cartesian coordinates corresponding to the total distance from the left end of the chromosome to that gene measured physically and genetically. The rate of recombination is thus given by the slope of the line connecting two points. This is a very versatile method, allowing multiple genetic maps to be displayed on the same axes, and lets the eye appraise the effects of uncertainties in the measurements very readily. For example, the error in the slope of each interval will depend only on the interval measurement, but the error in the mean slope over many intervals will depend on the sum of errors across the region
Assigning genetic coordinates.
To generate the data, first a subset of cloned markers were chosen which had sufficiently accurate 2-factor or 3-factor data between adjacent pairs, using the CGC genetic map data in acedb. In general, this means 2-factor distances scored from over 1000 F1 s,or 3-factor distances scored from over 30 recombinants. Secondly, the genetic distances between genes were recalculated from these data
Assigning physical coordinates.
The 920201 version of the physical map has 26 contigs genetically aligned, and 1900 kb (in 19 contigs) not yet aligned. The 26 contigs require 20 joins for closure. Hence, each join yet to be made was assumed to span 1900/20 -100 kb, which seems a reasonable first approximation. Each band from a fingerprint was assumed to span 1.8 kb a. Sulston, p.c.; 2 bands per Hind III site)
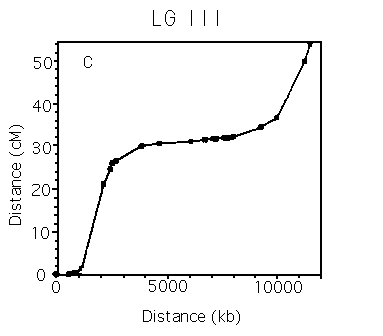
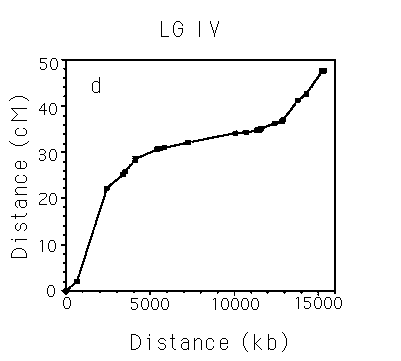
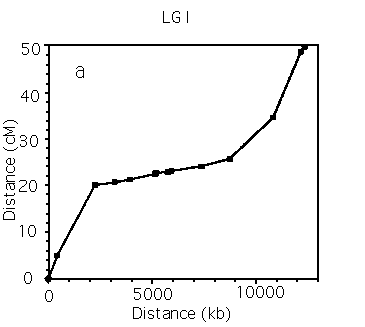
Analysis of Marey maps.
Autosomes. The Marey maps for the 5 autosomes are shown in panels a-e of the figure [See figures]. It is immediately apparent that they are all sigmoidal. This means that the rates of recombination are higher than average on the arms, and lower than average in the centers, which corresponds to earlier inferences (Greenwald et al., NAR 1987; Prasad and Baillie, Genomics 1989; Starr et al., Genome 1989). Also, each of the autosomes has a cluster that is clearly defined in physicogenetic terms. The terminal clusters on mL and XR also seem to resemble central clusters in that their rate of recombination decreases. For other terminal clusters, there are insufficient markers to determine whether they also behave like clusters. The X chromosome. There has been some debate over the years as to whether LG X contains a cluster. The Marey map for the X [See Figure 6] dearly shows that it does not contain a physicogenetic duster analogous to those seen for the autosomes - the X (apart from the terminal cluster) is linear. Panel f [See figure 6] also shows on the same axes data derived solely from STS mapping of stPs, which is consistent with a clusterless X (data from Williams et al., Genetics in press). Clusters. A comparison of the clusters [SeeFigure 7] shows that most of the autosomes are remarkably similar in both genetic and physical length. Also, each of the sub-intervals has roughly the same rate of recombination as the whole cluster (i.e. the clusters are linear), implying that the metric (no. of kb/cM) is relatively constant over the length of the cluster. The mean metrics for the autosomal clusters range from 1300 to 1900, but given the discussion of cumulative error above, it is reasonable to say that each of the clusters has a metric of roughly 1500. This can be seen more convincingly if the clusters are arranged in tandem [See Figure 8]. This value is clearly different from the central part of the X, which has a metric of about 450 [See Figure 7]. Thus it seems that all of the autosomal dusters have similar properties. Arms. In contrast, the different autosomal arms seem to have quite variable properties [See Figure 9]. The arms range from a mean of 90 kb/cM (IIIL, excluding the terminal cluster) to 370 kb/cM (VR). The arms vary widely in both genetic (12-30 cM) and physical (3-9 Mb) length.