Worm Breeder's Gazette 12(2): 94 (January 1, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
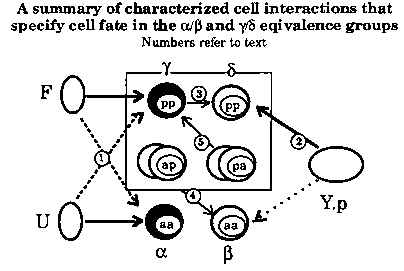
Cell fate determination in the B lineage of the male tail is due, in part, to cell interactions. Two equivalence groups, alpha/ß (B.a(l/r)aa) and gamma/delta (B.a(l/r)pp) are evident from the wild-type cell lineage [Sulston & Horvitz, Dev Biol 82:41-55 (1977)]. For instance, in the alpha/ß pair, either B.alaa or B.araa (labeled aa in the figure) can adopt a position anterior to the other. The cell that adopts the anterior position will adopt a fate (i.e., undergo an a lineage), and the posterior cell will adopt ,B fate. Thus, although the cells in each pair have equivalent potential, they adopt different fates depending on their relative positions. This abstract summarizes results of cell ablation experiments that begin to characterize cell fate specification in these two equivalence groups.
1) F and U promote anterior (a and y) fates.
Ablation of the male specific blast cells F and U in late L1 males results in abnormal a (7/7 animals) and y (5/7) lineages. Specifically, presumptive alpha's often behave like ß's, and presumptive gamma's undergo abnormal truncated lineages. lin-3 , let-23 , let-60 ,and lin-45 ,genes identified for their role in hermaphrodite vulval induction, also act in these male tail equivalence groups. Mutations that reduce wild-type function of these genes result in the same lineage defects seen in F-U- animals (see WBG 11.2, p. 103). In addition, the F/U signal is redundant. Lineages in F- or U- animals are generally wild-type (F-: 0/4 a abn; 1/4 alpha abn. U-:V4 gamma abn; 0/4 y abn.).
2) Y.p promotes posterior (ß and delta) fates.
Ablation of Y.p in late L1 males results in abnormal delta (6/7) and occasionally abnormal ß (2/7) lineages. Specifically, presumptive delta' s undergo an extra round of division. Y.p may play a minor role in providing posterior information for the alpha/ß pair. However, it appears to be neither necessary nor sufficient to specify ß fate (see section 4).
3) lin-12 acts in the gamma/delta equivalence group?
The role of lin-12 in the gamma/delta equivalence group shows similarities to its role in the hermaphrodite vulva. lin-12 (0)mutations result in transformation of delta to gamma [Greenwald et al., Cell 34:435-444(1983)]. Although lin-12 (d)mutations result in no gamma/delta phenotype on their own, ablation of F and U in lin-12 (d)animals results in full transformations of gamma to delta (3/3 transformed). Thus, lin-12 specifies delta fate, but the F/U signal can override lin-12 (d)and induce a normal gamma. In contrast, there is currently no evidence that lin-12 plays any direct role in alpha/ß cell fate specification.
4) Interactions between other B.a progeny and the alpha/ß equivalence group. B.a progeny play a significant role in specification of ß fate. Ablation of all B.a progeny except B.a(l/r)aa (i.e., B.a(l/r)pp-, B.a(l/r)pa-, and B.a(l/r)apin late L2 )results in both B.alaa and B.araa adopting alpha fate (4/5 both alpha). However, the presence of any left/right pair of B.a progeny is sufficient to correctly specify ß fate. In other words, wild-type lineages for B.a(l/r)aa were observed in animals with B.a(l/r)p-(3/3 wt), B.a(l/r)pa-and B.a(l/r)ap (3/3 wt), or B.a(l/r)pp-and B.a(l/r)ap- (2/2 wt) ablations, although occasionally division axes were skewed.
5) Interactions between other B.a progeny and the gamma/delta equivalence group. Experiments to test the interactions between other B.a progeny and B.a(l/r)pp are incomplete. Nevertheless, it is evident that the fates of B.a(l/r)pp (or, possibly their progeny), like B.a(l/r)aa, depend on their B.a neighbors. Gamma/delta are wild type in B.a(l/r)a ablated animals (3/3 wt). However, ablation of B.a(l/r)pa and B.a(l/r)aa results in truncated y lineages (4/4 truncated). Perhaps even more curious, ablation of B.a(l/r)ap and B.a(l/r)pa results in three rounds of division with abnormal axes in both presumptive gamma and delta (2/2 abnormal). It is not likely that this is a transformation of delta to gamma fate. One possible explanation is that B.a(l/r)aa (or their progeny) can actively promote cell division in B.a(l/r)pp (or progeny). In unablated animals, this activity is blocked by B.a(l/r)pa and B.a(l/r)ap. Additional experiments may help clarify the roles of B.a neighbors in specification of gamma and delta fate.