Worm Breeder's Gazette 12(2): 74 (January 1, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
The lin-26 gene, which encodes a zinc-finger protein that might act as a hypodermal differentiation factor, is defined by a single allele, n156 (Ferguson and Horvitz, Genetics 110, 17-72, 1985; Labouesse and Horvitz, 1991 C. elegans Meeting). n156 /Dfheterozygotes are inviable, which presumably accounts for the failure of non-complementation screens to identify additional lin-26 alleles. We are attempting to develop a different approach to isolate new lin-26 alleles based upon the availability of the cloned lin-26 gene. At the same time we would like to isolate mutations affecting the transcription unit overlapping with and related to lin-26 on its left side (see Labouesse and Horvitz, 1991 C. elegans Meeting).
The principle of our strategy is to use a cosmid that complements n156 as a balancer to isolate lethal mutations mapping to the 40 kb region represented by the cosmid. To test the feasibility of this method, we performed a small-scale experiment that has allowed us to recover one lethal mutation mapping a few kilobases from lin-26 .We constructed a strain, MT6208 ,of genotype: dpy-10 ( e128 ) unc-4 ( e120 )/ mnC1 [ dpy-10 ( e128 ) unc-52 ( e444 )1;Ex[ dpy-10 (+) F31B8 ).We generated the extrachromosomal array by injecting a the 4 kb dpy-10 (+)plasmid pAL129 (at 100 ng/µl, kindly provided by J. Kramer) with the 40 kb cosmid F31B8 (at 20 ng/µl), which rescues n156 ,into the germline of e128 e1201 mnC1 [ e128 e444 ]animals, looking for non-Dpy animals. One stable line was selected that generated (1) wild-type animals (genotype e128 e1201 mnC1 ;Ex(+)), (2) Unc animals (genotype e128 e120 ;Ex(+)), (3) sterile Unc animals (genotype mnC1 ,Ex(+)), and upon segregation of the array, (4) Dpy animals (genotype e128 e120 ),(5) Dpy Unc animals (genotype e128 e120 ),(6) sterile Dpy Unc animals (genotype mnC1 ).This strain is useful for the isolation of mutations carried by the e128 e120 chromosome, but not by the mnC1 chromosome as mnC1 suppresses recombination.
We reasoned that after mutagenesis a lethal lin-26 mutation, as well as any other lethal mutation rescued by the cosmid F31B8 ,should manifest itself as follows: a strain carrying such a lethal mutation on the e128 e120 chromosome should be (1) viable when the array is present (Uncs should be segregated) and (2) inviable when the array is absent (Dpy Uncs should be absent). We have thus mutagenized MT6208 animals with ethyl methanesulfonate and looked in a clonal screen for F1 animals that would not segregate Dpy Uncs but do segregate Dpys (suggesting that the array can be lost in the progeny of that particular F1 )and Uncs. Only F1 animals of P0 animals that segregated Dpys were examined, since if an animal did not segregate the array its progeny would not do so either.
To date we have examined the progeny of 2,727 F1 animals, which corresponds to 1,714 genomes: (1) 328 had a very low brood size and were not considered further; (2) 685 did not segregate Dpys (suggesting that the array had been stabilized) and were not considered further; (3) 223 did not segregate Uncs and Dpy Uncs but segregated Dpys (suggesting that a let mutation mapping to the part of LGII covered by mnC1 had been isolated); (4) 1,490 segregated Dpys, Dpy Uncs and Uncs; and (5) 1 segregated Dpys and Uncs but no Dpy Uncs and was assigned the allele number n2412 .With experience, we are decreasing the proportion of animals of the first two classes, by avoiding animals carrying too few eggs and animals fully rescued for the Dpy phenotype (animals with an intermediate Dpy phenotype generally segregate the array).
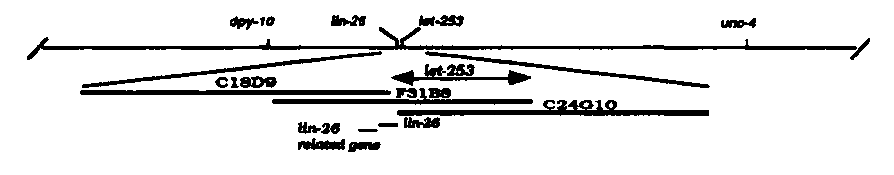
Further study of n2412 has shown that it complements n156 but does not complement let-253 ( mn181 ),a let mutation previously isolated by Bob Herman and his collaborators (Sigurdson et al., Genetics 108, 331-345,1984). At 20°C, both mn181 and n2412 do not truly cause lethality (although some embryos and larvae die) but rather a very slow growth, since most let-253 animals reach adulthood (within a week or more) and lay eggs. Preliminary examination of the animals under Nomarski optics shows that in let-253 animals most intestinal nuclei fail to divide during the L1 stage. Coupled with the observation that let-253 animals are severely constipated. The very slow growth phenotype might reflect a defect in intestinal cells function. In a few animals, the gonad also failed to develop on one or both sides. The cosmid F31B8 can rescue n2412 and mn181 animals. We have shown that the cosmid C18D9 does not rescue mn181 and have narrowed the region of F31B8 able to rescue mn181 to an 18 kb region of overlap between F31B8 and C24G10 (see Figure). This region maps to the right of lin-26 ,suggesting that let-253 does not correspond to the transcription unit overlapping with and related to lin-26 on its left side. We suggest the genetic order: dpy-10 , lin-26 , let-253 , unc-4 (see Figure).
Our approach could also be used to seek mutations in genes that have been defined by molecular biology rather than by genetic methods. It provides an alternative to PCR-based technologies (see Rushforth et. al., 1991 C. elegans Meeting) for the isolation of mutations within physically mapped genes for which a phenotype can be guessed based on protein sequence and domains of expression.
[See Figure 1]