Worm Breeder's Gazette 12(2): 71 (January 1, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
We are characterizing the myo-2 enhancer to identify factors controlling gene expression in pharyngeal muscle. To assay enhancer activity, DNA fragments cloned upstream of a lacZ fusion to the myo-3 or glp-1 promoter are tested for the ability to induce ß-gal in pharyngeal muscle.
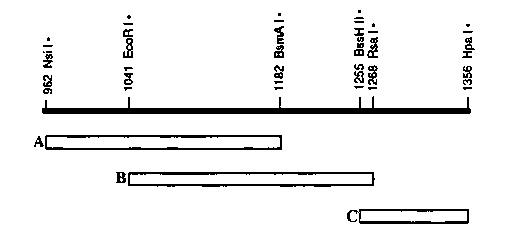
The myo-2 enhancer is pharyngeal muscle-specific and is located within a 0.4kb NsiI-HpaI fragment approximately 200bp upstream of the transcriptional start site. It can be divided into 3 fragments (A, B, and C) which are independently incapable of inducing pharyngeal expression from a myo-3 ::lacZfusion. However, any combination of two or more of these fragments (including duplications of any fragment) functions as an enhancer.
[See Figure 1]
As a working model, we propose fragments A, B, and C each contain a discrete enhancer element, and two or more elements are required for transcriptional enhancement. These elements are not functionally equivalent, however. We observe distinct patterns of pharyngeal muscle cell types expressing ß-gal when the entire myo-2 enhancer or the duplicated enhancer elements are assayed upstream of a glp-1 ::lacZfusion. The intact myo-2 enhancer induces expression in the major pharyngeal muscle cell groups ( m3 - m7 ),but not in the minor cell groups ( m1 and m2 ).Although the duplicated B element induces expression in a similar pattern of cells, ß-gal expression appears reduced in m5 and absent in m6 .In contrast, the duplicated C element gives expression in all the pharyngeal muscle cell types, including m1 and m2 .Finally, the duplicated A element is unable to enhance transcription from the glp-1 ::lacZfusion.
These results suggest the myo-2 enhancer elements contain binding sites for different transcriptional activators (or repressors), and the factors binding the B and C elements may be present in different sets of pharyngeal muscle cells. Furthermore, the observation that a functional enhancer requires multiple enhancer elements suggests the combination of factors present in any pharyngeal muscle cell type might determine the activity of the myo-2 enhancer. Consistent with this hypothesis, an enhancer containing the B and C elements is most active only in cells which express from both the duplicated B element and the duplicated C element ( m3 , m4 , m5 ,and m7 ).
Following protocols from Ginny Stroeher and Jim McGhee, we have begun examining worm nuclear extracts for proteins binding the myo-2 enhancer. Multiple retarded complexes are seen using the B element or the C element as a probe in gel retardation assays. These binding activities are specific for either the B or the C element. We are currently defining these binding sites to determine their role in myo-2 expression.