Worm Breeder's Gazette 12(2): 61 (January 1, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
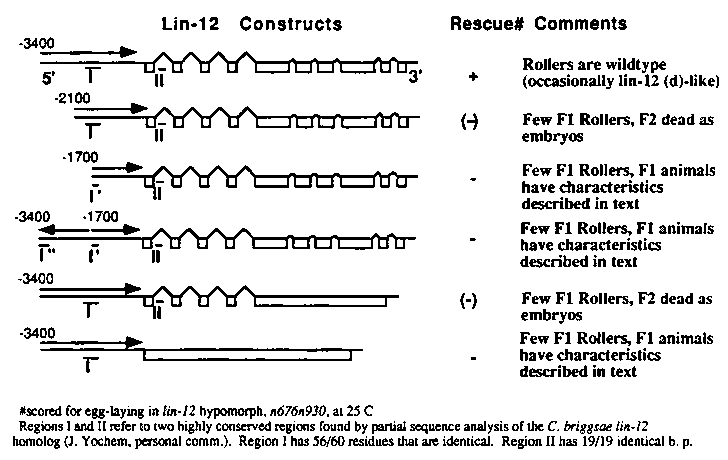
We are currently involved in defining sequences that may be relevant to lin-12 regulation. We have determined by primer extension and reverse PCR that the lin-12 message is trans-spliced to SL1 at a splice acceptor site 83 nucleotides upstream of the start of translation. DNA containing the coding region, 3.4 kilobase pairs (kb) of upstream sequence and .6 kb of downstream sequence is able to rescue the egg-laying defective phenotype of a lin-12 hypomorphic allele as well as the sterility of lin-12 null mutations.
In attempts to define the minimum genomic sequence needed for correct regulation we have constructed plasmids containing less 5' flanking region and fewer intron sequences. These constructs were injected at 1 µg/ml along with the dominant rol-6 marker (Mello et. al., EMBO J. 10: 3959-3970, 1991) at 100 µg/ml into the temperature sensitive lin-12 hypomorph, n676 n930 .The F1 progeny were screened for the Roller phenotype and then scored for rescue of the egg-laying defective phenotype at the restrictive temperature.
[See Figure 1]
We have found that there are sequences both 5' (approximately 1.7kb upstream of the start of translation) and internal (in one or more of the introns) that are needed for proper regulation. When these sequences are missing or disrupted we are unable to obtain Rollers at a reasonable frequency. Instead most of these constructs cause some F1 progeny of injected hermaphrodites to display a novel phenotype. These F1 animals are runty and necrotic but some do seem to proceed through larval development to adulthood (although they are sterile even when they have formed a gonad); they have extra rows of alae (often 2 rows, sometimes 4); their heads are abnormal with bumps between the first and second bulb of the pharynx or just posterior to the second bulb of the pharynx (the alae proceed in a straight line along the side of the animal but split around the bump at the head and rejoin on the other side); they also often have a bump at the posterior lip of the anus. These characteristics are reminiscent of some phenotypes associated with lin-12 (0)or lag mutations (Lambie and Kimble, Development 112: 231-240,1991); however, we have not determined that the cause of these defects is due to cell fate transformations. We suspect these defects are due to lin-12 expression from the extrachromosomal array because injection of the 5' sequence directs expression of a lacZ reporter gene (Fire et. al., Gene 93: 189-198, 1990) but does not cause this phenotype while injection of the lin-12 cDNA (constructed by K. Fitzgerald) with the same amount of 5' flanking sequence does cause the phenotype.
The presence of these putative control regions is consistent with the initial sequence analysis of the C. briggsae lin-12 homolog and we are defining them more specifically by making chimeric genes between the C. elegans and C. briggsae lin-12 genes. In addition we are studying their effects on the expression pattern of lin-12 :lacZchimeras.