Worm Breeder's Gazette 12(2): 106 (January 1, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
We are interested in identifying genetic loci whose zygotic expression is required for body-wall muscle development during embryogenesis. We have begun to screen available chromosomal deficiencies by staining homozygous embryos with muscle specific antibodies. A similar screen of deficiencies was carried out by F. Storfer and W. Wood (WBG 10(2):52) using light microscopy to assay various functions including muscle contractions. Use of monoclonal antibodies to the different muscle components has two advantages: First, we can determine whether a given cell is of the body-wall or pharyngeal muscle class. Second, we should be able to assay for commitment to a muscle cell fate (e.g. high level myosin expression) in the absence of any muscle function.
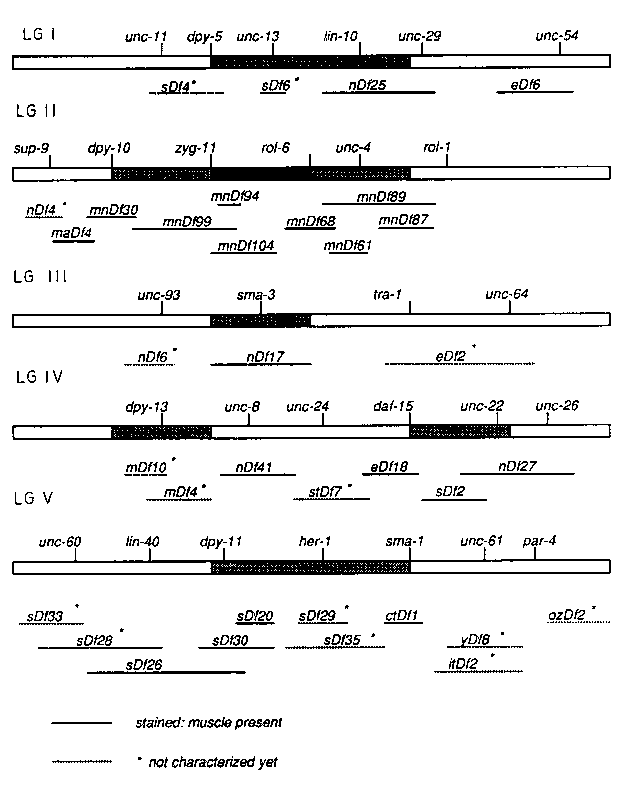
So far we have screened 20 deficiencies out of 35 deficiencies collected from CGC and other labs (see Fig.). Each strain carrying a deficiency was genetically manipulated to give rise to heterozygous hermaphrodite animals which reproducibly yield a large number of embryos. The total number of embryos laid and the number of unhatched embryos after 12 hours were counted in each case to determine what fraction of the arrested embryos are indeed deficiency homozygous. These arrested embryos were observed under polarized-light and nomarski microscope for their terminal phenotypes and stained with antibodies to muscle filament components such as the myo-3 and unc-54 gene products.
No deficiency that we have stained so far has shown total abolition of muscle staining. However, we have observed disorganized and abnormal muscle staining patterns in particular deficiencies (e.g. mnDf61 and mnDf99 ).In most cases, the terminal phenotypes of arrested embryos (e.g. twitching behavior and extent of morphogenesis) correlated with the degree of muscle development as judged by antibody staining. In some cases however, embryos arrested in morphogenesis earlier than comma stage showed normally developed muscle quadrant structures very similar to those seen in late stage wild type embryos.
The ctDf1 deficiency covering myo-3 gene on LGV (Fig.) showed only background levels of staining with antibody to myo-3 gene product but showed strong staining with antibody to unc-54 .Likewise the deficiency eDf6 covering unc-54 on LGI showed strong myo-3 staining but lacked unc-54 product. This suggests that only zygotic expression of myosins are detected in this immunostaining assay.
We will continue to screen available deficiencies which cover remaining regions on the genome. We would be grateful for contributions: i.e. any deficiency not listed in CGC genetic map and covering regions not covered by deficiencies shown in the Fig.
[See Figure 1]