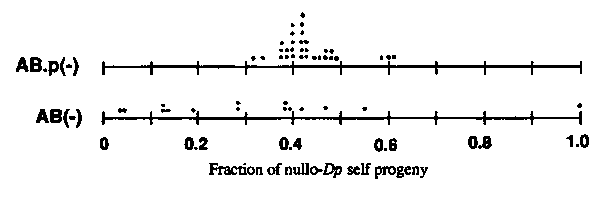
Worm Breeder's Gazette 12(2): 100 (January 1, 1992)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
The mechanism by which a free duplication is sometimes lost somatically is unknown. The experiments I here describe indicate that some fraction of losses--but not all--occur through nondisjunction. Self progeny of hermaphrodites of genotype dpy-1 ncl-1 unc-36 ; sD3 (III;f)[ dpy-1 (+) ncl-1 (+) unc-36 (+)]were screened for animals exhibiting an Unc-36 non-Dpy phenotype. The focus of unc-36 action is among descendants of AB.p (Kenyon 1986); the focus of dpy-1 action is diffusely distributed, apparently among hypodermal cells (E. Hedgecock, pers. comm.), which descend from both AB and P1 .A large proportion of the Unc-36 non-Dpy animals are generated by duplication loss at either AB or AB.p, which can be confirmed by scoring the Ncl phenotypes of appropriate cells (E. Hedgecock, pers. comm.). In this way I have identified AB(-) and AB.p(-) mosaics. I then scored their self progeny for the proportion that were nullo- sDp3 (Dpy Unc rather than wild-type). The presumption is that if an AB(-) mosaic is generated by nondisjunction of the duplication, then the two copies of sDp3 carried by P1 and its descendants, including the germ line, will lead to a reduced frequency of nullo- sDp3 gametes (and self progeny). The AB.p(-) mosaics provide a control, since their germlines should be unaffected. Assuming that the two duplications behave independently (no pairing at meiosis, for example), we would expect the frequency of nullo-Dp self progeny from a two-duplication germline to be the square of the frequency of nullo-Dp self progeny from a one-duplication germline. The results given below show that roughly a third to a half of the AB(-) mosaics gave a significantly lower frequency of nullo-Dp progeny than did the AB.p(-) mosaics. I take this as evidence for nondisjunction, and I shall try to confirm cytologically the presence of two duplications in the germline of particular animals.
One of the AB(-) mosaics apparently had no duplication in its germline, because it gave no duplication-bearing progeny. This animal could have been a consecutive mosaic; such mosaics, first identified by E. Hedgecock, suffer duplication loss at two or more consecutive divisions. Two of the Unc-36 non-Dpy animals picked in the above screen were in fact triple consecutive mosaics: each was P1 (-)AB.a(-) AB.pr(-). It seems very unlikely that the losses that occur in consecutive mosaics involve nondisjunction.
In our mosaic analysis of lin-15 ,E. Hedgecock and I found that about two-thirds of P1 (-)mosaics were Muv (or partially Muv) and one-third were wild-type. We concluded in that study that the focus of lin-15 action was in the hyp7 syncytial hypodermis, which derives from both P1 and AB. But if a P1 (-)mosaic were produced by nondisjunction, then the loss of lin-15 (+)from the P1 -derived hyp7 nuclei might be compensated for by the gain of an extra lin-15 (+)copy in the AB-derived hyp7 nuclei. This suggests that the non-Muv P1 (-)mosaics were produced by nondisjunction of the free duplication ( mnDp14 ,in this case) and the other P1 (-)mosaics were produced by straight duplication loss.
[See Figure 1]