Worm Breeder's Gazette 12(1): 36 (September 1, 1991)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
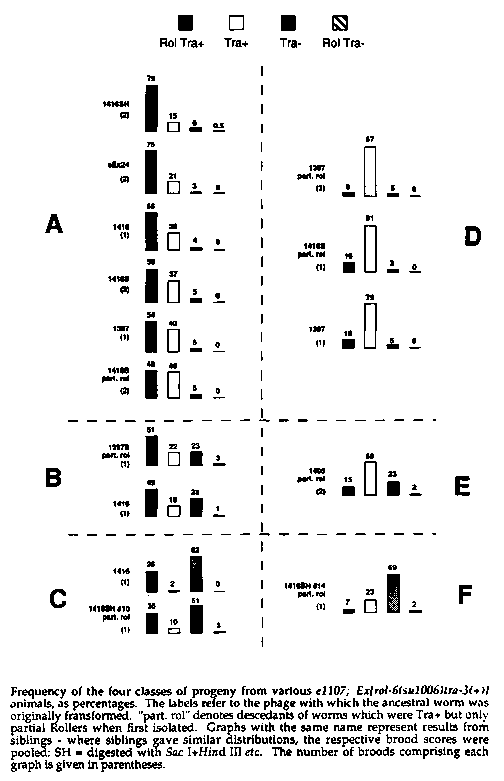
Early on I noticed that some e1107 ; eEx24 animals have Tra-3 F1 s.Two broods were fully scored, and the frequency of Tra-3 animals was found to be 3% (see figure). The failure of complete maternal rescue suggests that the diffusion range of the tra-3 product is low, by the following argument. In the distal part of the gonad, germ cells are kept dividing mitotically by a signal mediated by the glp-1 gene product (Austin and Kimble, Cell 1987). As they move away from this signal, they enter meiosis. Hence array loss from a mitotic germ cell will lead to a clonal proliferation of tra-3 (0)nuclei which would tend to be situated together on the rachis. A limited diffusion range would lead to Tra-3 (0)oocytes, which would appear in the F1 .In the heterozygote mother ( e1107 /+),complete rescue is seen because nuclei remain diploid until after fertilization.
Examining the pattern of phenotypes produced from other arrays showed that some had a much higher percentage of Tra-3 F1 sthan eEx24 .In addition, some seemed to produce very few Rol F1 s.To examine this further, I scored the phenotypes for several broods of a number of different arrays (see figure). Several things became apparent. First, two siblings can produce broods with very different characteristics (e.g. 1416 and 1397 siblings). Secondly, any given brood can be quite cleanly placed in one of six classes, i.e. those defined by the independent assortment of whether there are few, an intermediate number, or many Tra-3 F1 sand whether there are many or few Rol Tra+ (classes A to F in figure). The frequency of rolling tra-3 -rescued F1 sis presumably related to the degree of mitotic and meiotic array loss in the germline of the parent, and is simply a measure of transmission rate. This can vary between siblings, but apparently is in only one of two states (low or high). The three degrees of Tra-3 F1 production, independent of the frequency of Rol Tra+ production, presumably represent different levels of expression of tra-3 .Again, this can vary between siblings, and is in one of a few relatively discrete states. This suggests that the various classes do not reflect properties of the arrays as such, but rather are markers of their fates in the animal. The tra-3 product is unlikely to be secreted (see other Barnes abstract elsewhere in this issue), so the source of all tra-3 product is likely to be from within the syncytium.
Class A broods are then explained as for eEx24 above, with transmission occurring through both sperm and oocytes. Class C broods have a fair degree of transmission, but little maternal rescue, which suggests transmission is occurring principally through the sperm. Class D broods have little transmission, but potent maternal rescue. If one presumes that for some reason transmission through oocytes is less successful than through sperm, this class could be interpreted as having largely lost the array from the sperm. Class F would then be like class D, except that even fewer array copies would be in the female germ line, such that the threshold of rescue was infrequently reached. Since events in the two gonad arms are expected to be independent, classes B and E could represent arms of classes A+C and D+F, respectively. In fact, the 1416S part. rol results (class A) resemble an A+D result.
The final observation to be made from these results is that there is a very low, but real, frequency of Rol Tra-3 - F1 s. rol-6 encodes a cuticular collagen, which presumably is required in reasonable amounts to produce rolling; it is known from other work (see e.g. Kondo et al., Mol. Cell. Biol. 1988) that very little tra-3 expression is required to rescue mutants. These unusual F1 smust have received the array because rol-6 ( su1006 ),the dominant allele used, has no maternal effect (at least in single copy); since tra-3 does have a maternal effect, the oocytes that gave rise to these zygotes must have been Tra-3 -.Therefore, in some as yet unsatisfactorily explainable fashion, tra-3 expression from the array must have been selectively shut off in these oocytes and resultant zygotes, either by DNA loss or regulation.
[See Figure 1]