Worm Breeder's Gazette 12(1): 29 (September 1, 1991)
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
We chose C. elegans as a model to obtain clues as to the physiological function of P-glycoproteins (see accompanying paper) in a relatively simple multicellular organism. As a first step in this analysis, we determined mRNA levels of three C. elegans P-gp genes (cepgpA,-B & -C) during development (with RNase-protection assays; internal standard: actin-IV mRNA), and the tissue-specific expression of two of them (cepgpA & -C) by transformation of nematodes with lacZ-fusion constructs. The three genes were expressed throughout the life cycle of the nematodes. cepgpB mRNA levels were invariable. In contrast, cepgpA and cepgpC were found to be developmentally regulated with maximum steady state mRNA levels in early (cepgpA) and late larval development (cepgpC). DaD-lacZ fusion constructs: We made three constructs starting from Andrew Fire's pPD26 .50:
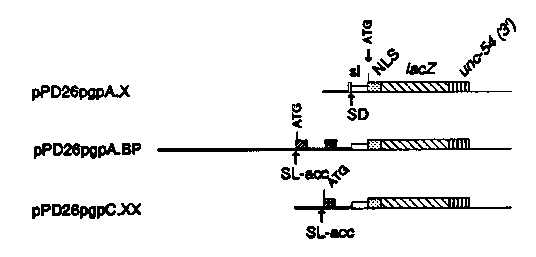
[See Figure 1]
pPD26 pgpA.X:The glp-1 promoter was removed (except 15 nucleotides 5' adjacent to the synthetic intron, providing a splice donor (SD)). A 460 bp fragment from cepgpA, containing the 5' flanking sequence of the first exon but not its spliced leader acceptor (SL-acc), was inserted.
pPD26 pgpA.BP:The glp-1 promoter and part of the synthetic intron (SI) was removed and replaced by a 4kb fragment containing 2.9 kb of 5' flanking sequence of cepgpA, its first two exons and part of the second intron.
pPD26 pgpC.XX:Analogous to pPD26 pgpA.BP,but containing a 1200bp fragment with 1 kb of 5' flanking sequence of cepgpC, its first exon and part of the first intron.
These constructs were co-injected with pRF4 (Jim Kramer) into N2 animals. rol-6 transmitting lines were established with each construct, and X-GAL-stained according to Andrew Fire's protocol (WBG11:2). Results: Independent lines from all three constructs had similar staining patterns, although the intensity of the stain differed somewhat, even between lines transformed with the same construct. In almost all animals the staining started in the region proximal of the anus and just distal of the pharynx. After continued incubation the staining increased in intensity and progressed towards the middle portion of the animals. No staining was observed proximal of the pharyngeal-intestinal valve and distal of the anus. Some animals showed a clear nuclear staining pattern (very similar with all the three constructs) with little or no diffuse tissue staining, suggesting that the nuclear targeting signal (NLS), present in all constructs, was functional. We interpret the nuclear pattern as indicative of exclusive expression in intestinal cells. Surprisingly, the staining was most intense in L1 /L2stage animals with pPD26 pgpA.Xand pPD26 pgpA.BP,whereas animals transformed with pPD26 pgpC.XXalso showed staining in later stages (in agreement with RNase protection analysis of stage specific expression). Apparently, the fusion protein is not very stable.
Expression of cepgpA and cepgpC in intestinal cells (and overexpression upon mutagenesis and drug selection; see accompanying paper) suggests that P-gp's may protect C. elegans from ingested toxins. This pattern is in good agreement with that of some mammalian P-gp isoforms, that are most abundantly expressed in the epithelia of the gut and that of excretory organs.
(We thank Ian Hope, University of Leeds, U.K., for helpful advice in the interpretation of the staining pattern)