Worm Breeder's Gazette 11(5): 96
These abstracts should not be cited in bibliographies. Material contained herein should be treated as personal communication and should be cited as such only with the consent of the author.
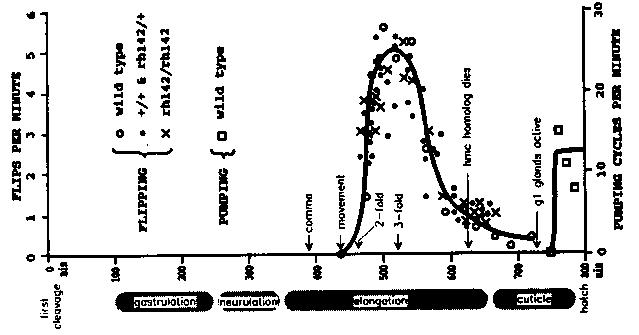
unc-104 mutations cause a generalized loss of chemical synapses, including neuromuscular junctions, and disrupt neural regulation of behaviors including locomotion, feeding, and defecation (1). unc-104 is a kinesin-related protein and, by inference, functions as a microtubule-based motor (2). Based on behavioral and anatomical phenotypes, we suggest this motor is neuron-specific and used for anterograde transport of vesicles to presynaptic active zones but not for axonal growth. We examined embryonic and molting behaviors in sublethal null rh142 or severe viable rh43 mutants, respectively. FLIPPING - Body muscle contractions are required during embryonic elongation, possibly to ensure that myofilaments align along the longitudinal axis of the hypodermis (3,4). These contractions begin at 430 minutes, shortly after the onset of embryonic elongation (5). Although movements are small and localized at first, by 460 minutes (2- fold) all dorsal or ventral muscles contract synchronously and vigorously. These contractions alternate regularly between dorsal and ventral muscles (up to 5 per minute) and continue throughout elongation. Because the embryo is folded within its rigid eggshell, each longitudinal contraction creates torque which flips the embryo 180 about its anteroposterior axis. At the onset of cuticle synthesis, flipping declines abruptly and the larval locomotor pattern (spatially & temporally alternating contractions) emerges. In rh142 embryos, early body muscle movements closely resemble wild type and rates of flipping are comparable throughout elongation (see Figure). Flipping declines normally but the larval pattern never emerges. Instead animals often assume coiled postures for minutes at a time. These embryonic contractions may spread via gap junctions between arms of neighboring body muscles (6). The contraction rate could be controlled autonomously by muscle cells. Alternatively, any of three mesoglia (hmc, hmc homolog, GLR), which form gap junctions with body muscles in the head, could act as pacemakers (7). [See Figure 1] HATCHING - About 30 minutes before hatch, pharyngeal muscles begin contracting at 10-15 cycles per minute; each pumping cycle comprises simultaneous contraction of the corpus and terminal bulb, followed by contraction of the isthmus (5). The intestine, initially collapsed, quickly becomes distended with ingested fluid. This extraembryonic fluid appears to recirculate through the alimentary tract by a cycle of ingestion, and presumably release through the rectum, that continues until hatch. Eggshells invariably rupture within 20-30 minutes after the onset of pumping. In most rh142 embryos, the pharynx never contracts or quivers only briefly, and the intestine remained undistended. In some embryos, one or more complete pumping cycles occur and the intestine becomes slightly distended. Generally, hatch is delayed by 30-120 minutes and some 10% of mutant larvae fail ever to rupture their eggshell. The purpose of embryonic pumping is unknown. The extraembryonic fluid could acquire enzymes necessary for digesting the eggshell while recirculating through the pharynx and intestine. Possible sources for these enzymes are the pharyngeal gl glands and the intestine itself (5, 8). Hatch-defective mutants of the hch-1 gene may identify a protease for digesting the outermost shell (10). MOLTING - Pharyngeal gl glands are exocrine cells with axon-like processes connecting cell bodies in the terminal bulb to active zones of exocytosis at the anterior (g1P) or posterior (g1AL/g1AR) limit of the corpus (9). These cells are continuously active during feeding but have a novel cycle of secretion during molts (8). During the lethargus, large secretory granules accumulate in the gland cell bodies. About 1 hr before ecdysis, granules begin entering the g1P process but not the shorter g1P processes. Milling irregularly, they gradually fill the proximal segment (isthmus) but do not enter the distal segment (corpus). By 30 minutes before ecdysis, the proximal segment of the g1P process and both g1A cell bodies are densely packed with granules. Abruptly, granules in all three cells begin rapid, steady anterograde translocation (ca. 1 micron/second) to the active zones where they are released within a few minutes. Possibly these secretions help loosen and weaken the old cuticle where it must tear apart at ecdysis (8). In rh43 larvae, lethargus is generally longer than normal and the timing of granule synthesis and transport is poorly regulated. However, each molt concludes with a rapid, steady anterograde translocation of secretory granules. These mutants suggest that distinct vesicle populations, with separate molecular motors, are used for axonal growth, synaptogenesis, and exocrine functions. In particular, secretory granules are likely translocated by an anterograde motor distinct from unc-104.